
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Дипломная работа: Содержание продуктов перекисного окисления липидов в плазме крови беременных женщин
Дипломная работа: Содержание продуктов перекисного окисления липидов в плазме крови беременных женщин
Федеральное агентство по образованию
ФГОУ ВПО Сибирский федеральный университет
Институт фундаментальной биологии и биотехнологии
Кафедра биохимии и физиологии человека и животных
В.С. Выборова
студентка V курса
Содержание продуктов перекисного окисления липидов в плазме крови беременных женщин
(дипломная работа)
Допустить к защите:
Зав. кафедрой, д.м.н., проф. А.А. Савченко
Научный руководитель:
Канд. биол. наук, проф.
Н.М. Титова
Красноярск 2008
Содержание
Введение
Глава 1. Обзор литературы
1.1. Активные формы кислорода классификация и свойства
1.2. Перекисное окисление липидов
1.3. Антиоксидантная система
Глава 2. Материалы и методы
2.1 Объект исследования
2.2. Определение содержания малонового диальдегида
2.3. Определение содержания диеновых коньюгатов
2.4. Статистическая обработка результатов
Глава 3. Результаты исследований и обсуждение
3.1. Содержание продуктов перекисного окисления липидов в плазме крови небеременных женщин
3.2. Содержание диеновых коньюгатов и малонового диальдегида и в плазме крови женщин в разные периоды беременности
Выводы
Список литературы
Summary
В последние годы широко обсуждается роль активных форм кислорода (АФК) и инициируемых ими свободнорадикальных процессов при различных патологических процессах, а так же при беременности. В нормальных условиях активность этих процессов находится на невысоком уровне, но при стрессовых ситуациях происходит усиленное образование АФК, под действием которых происходит избыточная и неконтролируемая активация процессов перекисного окисления липидов (ПОЛ), что в конечном итоге может привести к патологическому состоянию, которое сопровождается дисбалансом ферментативных и неферментативных компонентов системы антиоксидантной защиты. Физиологическая беременность может сопровождаться существенными изменениями в про/антиоксидантном статусе.
Характерным проявлением окислительного стресса является интенсификация процессов перекисного окисления липидов, индикатором которой служит увеличение содержания хотя бы одного из его продуктов. Данные о содержании продуктов ПОЛ в биологических объектах могут нести в себе информацию о глубине и степени патологического процесса. В качестве количественных маркеров наиболее часто используются такие интермедиаты ПОЛ, как диеновые конъюгаты (ДК), а также один из его конечных продуктов малоновый диальдегид (МДА).
Цель данной работы определение содержания ДК и МДА в плазме крови у женщин в разные периоды беременности.
В задачи работы входило:
1. Определить содержание продуктов перекисного окисления липидов в плазме крови небеременных женщин.
2. Определить содержание ДК в плазме крови женщин в динамике беременности.
3. Определить содержания МДА в плазме крови женщин в динамике беременности.
Работа выполнена на базе кафедры биохимии и физиологии человека и животных Института фундаментальной биологии и биотехнологии Сибирского Федерального университета и Хакасского республиканского центра планирования семьи.
Глава 1. Обзор литературы.
1.1. Активные формы кислорода – классификация и свойства.
Термин активные формы кислорода (АФК) объединяет целый ряд образующихся в организме промежуточных и побочных продуктов восстановления молекул кислорода, таких как супероксидный (О2-), гидроксильный (НО·), пергидроксильный (НО2∙), пероксильный (RO2∙) и алкоксильный (RO∙) радикалы, оксид азота (NO∙), пероксинитрит (ONOO-), гипохлорит (HOCl), перекись водорода (H2O2) и др. Помимо продуктов восстановления О2, к АФК относят также озон (О3) и синглетный кислород 1О2, то есть кислород, находящийся в возбужденном (синглетном) состоянии [Владимиров, 1998; Осипов, Азизова, Владимиров, 2003]. Все АФК, кроме синглетного кислорода, представляют собой разные химические соединения: молекулы (Н2О2), свободные радикалы (ОН, НО2), ион-радикалы (О2-). Поэтому термин активные формы кислорода следует считать собирательным, он подчеркивает высокую реакционную способность промежуточных продуктов восстановления молекулы кислорода, кислородных радикалов и их прекурсоров.
Радикалы кислорода образуются в ходе процессов, связанных с транспортом электронов по дыхательной цепи. В обычных условиях на генерацию АФК расходуется от 1 до 5% потребляемого кислорода. Однако эта величина может существенно возрастать при изменении кислородного бюджета организма – при гипероксии или гипоксии. В результате последовательного восстановления молекулярного кислорода происходит образование супероксидного аниона, перекиси водорода, гидроксильного радикала.
Супероксидный анион-радикал (О2-). Среди кислородных свободных радикалов ему отводят наиболее значительную роль, так как считается, что именно он является родоначальником многих других активных форм кислорода. Супероксидный радикал (СОР) образуется при присоединении одного электрона к молекуле кислорода в основном состоянии [Chen, Warden, Stenken, 2004]. Химическая активность О2- в значительной степени зависит от физико-химического состояния окружающей его клеточной или внеклеточной среды. В водных растворах О2- способен окислять аскорбиновую кислоту, адреналин и тиоловые соединения, выступая как слабый окислитель [Хавинсон, Баринов, Арутюнян 2003]. Значительно более выражены восстановительные свойства супероксидного радикала. В присутствии ионов негемового железа СОР достаточно активно восстанавливает его из трехвалентного в двухвалентное состояние. Это свойство СОР чрезвычайно важно, поскольку двухвалентное железо играет большую роль в образовании агрессивных липидных и гидроксильных радикалов [Владимиров, 1998]. Супероксидный радикал также может восстанавливать содержащие трехвалентное железо комплексы (цитохром с, ферри-ЭДТА) и нитросиний тетразолий [Auchere, Rusnak 2005].
Образование супероксида в организме в основном происходит при работе митохондриальной и микросомальной цепей переноса электронов, в результате «утечки» электронов с восстановленных элементов этих цепей на молекулярный кислород [Осипов, Азизова, Владимиров, 2003], при активации фагоцитирующих клеток крови и тканевых макрофагов [Владимиров, 1998], в ходе энзиматических реакций при действии, так называемых «перекись продуцирующих ферментов», моно-и диаминооксидаз, моно-и диоксигеназ, при окислении гемоглобина и миоглобина, а также любых, склонных к аутоокислению биомолекул: аскорбиновой кислоты, восстановленного глутатиона, биогенных аминов [Хавинсон, Баринов, Арутюнян, 2003].
Супероксиданион-радикал пусковое звено каскада радикальных реакций, приводящих к возникновению большинства АФК и продуктов перекисного окисления липидов. Он участвует в синтезе хемотаксических пептидов, усиливает митогенстимулированную пролиферацию лимфоцитов, ингибирует действие эндотелиального фактора расслабления, может повреждать мембраны эритроцитов, ингибировать Са-АТФазу, синтез РНК и белка эндотелиальных клеток, окислять белки сыворотки, в тоже время его непосредственная цитотоксичность невелика [Воейков, 2004].
Для регуляции уровня О2- в клетках служит высокоспецифичный фермент-антиоксидант – супероксиддисмутаза (СОД), которая обладает существенной способностью ускорять реакции дисмутации радикала с образованием молекул перекиси водорода и кислорода.[Okado-Matsumoto, Fridovich, 2003]
О2- + О2- + 2Н+ → Н2О2 + О2
Пероксид водорода. Н2О2 не является свободным радикалом. Образование О2- в любой биологической модельной системе сопровождается накоплением Н2О2, образующимся в результате дисмутации (неферментативно или в присутствии супероксиддисмутазы) [Дубина, 2004]. Поэтому в организме повышение концентрации Н2О2 наблюдается при активации процессов, которые связаны с генерацией супероксидного радикала: при состояниях метаболического взрыва фагоцитирующих клеток; при усиленной деятельности митохондриальных и микросомальных электронтранспортных цепей; при повышении активности оксидазных ферментов.
Будучи стабильным продуктом восстановления кислорода, Н2О2 обладает свойствами слабого окислителя. Эти свойства проявляются, в частности, в присутствии ионов металлов с переменной валентностью в восстановленной форме [Осипов, Азизова, Владимиров, 2003], в результате чего образуется высокоактивный гидроксильный радикал:
Н2О2 + Fe2+ → Fe3+ + OH- + OH
В отсутствии каталазы и ионов металлов переменной валентности пероксид водорода довольно стабилен и вследствие своей незаряженной ковалентной структуры воспринимается клеткой, как молекула воды. Благодаря этому Н2О2 может легко проникать в клетки и ткани, при этом наличие нейтральных аддуктов пероксида водорода (например, гистидина) обеспечивает проникновение внутрь клеток даже в присутствии каталазы. Считается, что биологическая активность Н2О2 существенно зависит от её концентрации. Так, при низких (микромолярных) уровнях Н2О2 является относительно слабореактивной. Однако с ростом концентрации агрессивность пероксида водорода увеличивается и при достаточно высоком (миллимолярном) уровне Н2О2 обладает цитотоксическим действием и может вызывать гибель фибробластов и других типов клеток, включая гепатоциты и эндотелиальные клетки. В сублетальных концентрациях пероксид водорода существенно изменяет статус эндотелиальных клеток, что проявляется в ингибировании транспорта анионов через мембрану, увеличении внутриклеточной концентрации Са2+, активации фосфолипаз и фосфоинозитидного обмена, повреждает Сu,Zn-СОД, тем самым снижая антиоксидантную защиту клеток [Меньщикова с соавт., 2006].
Клетки млекопитающих достаточно устойчивы к воздействию пероксида водорода, благодаря наличию глутатионпероксидазной и каталазной ферментативных систем, первая из которых эффективно работает при малых концентрациях перекиси, вторая – при высоких.
Гидроксильный радикал (НО∙). Одноэлектронное восстановление Н2О2 приводит к образованию гидроксильных радикалов, обладающих чрезвычайно высокой реакционной способностью. Разложение Н2О2 в присутствии ионов двухвалентного железа является основным путем образования НО∙ (реакция Фентона) [Владимиров, Арчаков, 2003]:
H2O2 + Fe2+ → Fe3+ + OH- + HO∙
Другой путь образования гидроксильного радикала – это реакция разложения гипохлорита, которая также протекает с участием Fe2+ [Осипов, Якутова, Владимиров 2003; Якутова с соавт., 2004]:
НОCl + Fe2+ → HO + Cl- + Fe3+
Установлено, что образование гидроксильного радикала возможно при разложении гипохлорита также и железонезависимым путем
HOCl +O2- → HO + Cl- + O2-
Вследствие высокой химической активности время жизни ОН-радикалов в клетке составляет около 10-9 с, а расстояние, которое они успевают пройти за это время от места их образования, не превышает 100 нм. Таким образом, клеточная топография повреждающего действия ∙ОН-радикалов и, как следствие этого, характер эффекта повреждения будет зависеть от места их образования. Например, возникновение ОН-радикалов вблизи молекулы ДНК с высокой вероятностью приведет к модификации основания и взрыву одной из цепей ДНК [Kira, Sato, Inoue 2003].
Обладая наиболее высоким в живой природе редокс-потенциалом (Е0=+2.7В), и будучи вследствие этого чрезвычайно агрессивным, ∙ОН оказывает действие практически на любую биологическую молекулу. Но наибольший ущерб клетке наносят его реакции с ДНК, белками и полиненасыщенными жирными кислотами внутриклеточных и плазматических мембран, что определяет сильнейшее мутагенное и цитотоксическое действие гидроксильного радикала [Melov, 2003].
Важно отметить, что в организме нет специальных ферментативных систем, обладающих способностью инактивировать гидроксильный радикал. Низкомолекулярные соединения, такие как урацил, мочевая кислота, салицилаты, глюкоза, диметилсульфоксид, обладают способностью ингибировать ∙ОН-радикал только при достаточно высоких концентрациях [Зенков, Ланкин, Меньщикова, 2004]. Таким образом, при целом ряде патологических состояний, сопровождающихся избыточным образованием АФК и, соответственно, гидроксильного радикала, организм становится практически беззащитным перед повреждающим действием этого соединения. Предотвращение повреждений клеточных структур осуществляется только за счет снижения концентрации радикалов предшественников ОН, в частности, супероксиданион-радикала и пероксида водорода.
Синглетный кислород (1О2). Образуется при изменении спина одного из электронов π-орбитали в молекуле кислорода. Источником синглетного кислорода являются реакции фотосенсибилизированного окисления биологических субстратов [Осипов, Азизова, Владимиров, 2003]. При нефотохимических реакциях образование 1О2 возможно в результате неферментативной дисмутации супероксидных радикалов, протекающей с образованием перекиси водорода в присутствии ионов металлов с переменной валентностью:
О2- + О2- + Н2 → Н2О2 + 1О2
А также при взаимодействии некоторых сильных окислителей, например гипохлорита с Н2О2 и ферментативно – в реакциях восстановления цитохрома с [Maiorino, Zamburlini, Roveri, 2005]. В отличие от молекулы О2, синглетный кислород обладает высокой химической активностью, особенно по отношению к молекулам, содержащим участки повышенной электронной плотности (ненасыщенные жирные кислоты, ароматические аминокислоты, основания). Типичным для 1О2 являются реакции взаимодействия с двойной связью. Это свойство 1О2 особенно важно для инициирования перекисного окисления ненасыщенных липидов в биологических мембранах [Меньщикова с соавт., 2006]. Кроме того, синглетный кислород, как и гидроксильный радикал, вызывает окисления сульфгидрильных групп в белках, декарбоксилирует аминокислоты, расщепляет нуклеиновые кислоты [Eisenberg, Taylor, Guerrero 2002]. Энергичное образование 1О2 в клетке может приводить к её повреждению или даже к гибели [Осипов, Азизова, Владимиров, 2003].
Одним из наиболее эффективных гасителей синглетного кислорода в клетке является β-каротин, одна молекула которого способна потушить около 1000 его молекул прежде, чем он подвергнется окислительной деструкции [Владимиров, Арчаков, 2003].
Гипохлорит (НОСl). Хлорноватистая кислота – сильнейший окислитель, образуется в нейтрофилах при участии гем-содержащего цитоплазматического фермента миелопероксидазы [Klebanoff, 2006]. Миелопероксидаза окисляет ионы солей хлористоводородной кислоты, Сl- в присутствии Н2О2 в ходе реакции:
Н2О2 + Сl- + Н+ → НОСl + Н2О
НОСl не является свободным радикалом, но выступает как один из наиболее сильных окислителей.
НОСl атакует простейшие амины, сульфгидрильные группы в белках и хлорированные пуриновые основания в ДНК [Хавинсон c соавт., 2003]. НОСl может взаимодействовать с замещенными арил-аминами (например, с анилином, 1-нафтиламином и 1-нафтолом) даже при физиологических уровнях, образуя долгоживущие продукты, которые связываются с ДНК и являются генотоксичнымим для клеток человека [Осипов, Азизова, Владимиров, 2003].
1.2. Перекисное окисление липидов.
Все активные формы кислорода обладают высокой цитотоксичностью в отношении любых типов клеток и клеточных образований, что определяется их химической активностью. Можно выделить 4 наиболее вероятные мишени окислительной цитотоксической атаки АФК: индукция процессов ПОЛ в биологических мембранах, повреждение мембраносвязанных белков, инактивация ферментов и повреждение ДНК клеток.
Одним из важнейших следствий избыточного образования АФК является избыточная в этих условиях активация процессов ПОЛ [Барабой, 1991]. В нормальных условиях активность этих процессов находится на невысоком уровне, обеспечивающим протекание ряда физиологических процессов. Чрезмерная, патологически усиленная активация процессов ПОЛ под действием АФК приводит к необратимому изменению или повреждению мембранных структур, нарушению их проницаемости для ионов. Процессы ПОЛ можно условно подразделить на 3 последовательных этапа, или фазы развития: процесс зарождения цепей, процессы развития цепных реакций и обрыв цепей [Зенков, Ланкин, Меньщикова, 2004].
В биологических мембранах окислению подвергаются преимущественно полиненасыщенные жирные кислоты, входящие в состав фосфолипидов [Dix, Aikens, 2005]. На стадии инициирования под действием свободных радикалов О2, ионизирующей радиации, ультрафиолетового облучения и ряда химических веществ, относящихся к прооксидантам, происходит отрыв атома водорода в альфа-положении по отношению к двойной связи. Присутствие двойной связи в жирной кислоте (ЖК) ослабляет связь С-Н в смежных углеродных атомах и тем самым облегчает отщепление Н∙. Чем длиннее ненасыщенная боковая цепь кислоты жирного ряда, тем сильнее у неё склонность подвергнуться липидному окислению [Хавинсон, Баринов, Арутюнян, 2003]. Радикал с углеродом в центре претерпевает молекулярную перегруппировку с образование диена, содержащего сопряженные двойные связи, который в дальнейшем соединяется с О2 и образует радикал пероксида, способный отделить атом водорода от другой ЖК. Возникновение в результате этой реакции органических перекисей и нового радикала способствует продолжению окислительных реакций, приобретающих цепной характер [Зенков, Ланкин, Меньщикова, 2004].
Таким образом, перекисное окисление липидов представляет собой процесс, связанный с активацией кислорода, особенность которого заключается в том, что молекула О2 присоединяется к свободному радикалу [Meral, Tuncel, Surmen-Gur, 2000]:
О2 + L∙ → LO2
В результате получается новый пероксильный радикал органического соединения. В дальнейшем происходит взаимодействие этого радикала с новой молекулой органического соединения, в результате чего протекает процесс цепного ПОЛ.
LO2∙ + LH → LOOH + L
L∙ + O2 → LO2∙
Реакции перекисного окисления липидов указаны на рис. 1.
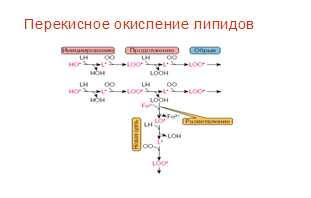
Рис.1. Реакции перекисного окисления липидов [Владимиров, 1998]
Считается, что образование перекисей липидов осуществляется двумя путями: неферментативным аскорбатзависимым (аскорбиновая кислота регенерирует ионы за счет обратного восстановления Fe3+ до Fe2+), активируемым металлами с переменной валентностью, и ферментативным (НАДФН-зависимым). По первому пути образование перекисей липидов происходит во всех мембранных структурах, а по второму преимущественно в эндоплазматическом ретикуломе [Gutteridge, 2005].
Ферментативное ПОЛ относится только к генерации липидных перекисей в активном центре фермента. Образованные при этом гидроперекиси и эндоперекиси являются стереоспецифическими и имеют важные биологические функции. В частности, это относится к циклооксигеназе и липооксигеназе.
Особенность неферментативного цепного окисления в биологических мембранах заключается в том, что оно практически не происходит в отсутствие металлов с переменной валентностью, в частности ионов двухвалентного железа [Владимиров, Арчаков, 2003]. Резко выраженное прооксидантное действие Fe2+ на ненасыщенные жирные кислоты (НЖК) в биологических мембранах обусловлено разложением продуктов реакции гидроперекисей, в результате которого в системе появляются новые свободные радикалы, инициирующие новые цепи окисления:
Fe2+ + LOOH → Fe3+ + OH + LO
При этом радикал LO вступает в дальнейшие реакции цепного окисления:
LO + LH → LOH + L
L + O2 → LO2
В последние годы появилось много данных, свидетельствующих, что ионы железа активируют процессы перекисного окисления также за счет того, что они участвуют в образовании гидроксильного радикала [Владимиров, 1998].
В ходе ПОЛ субстрат (полиненасыщенные ЖК) продолжают расходоваться, если только в этот процесс не вмешивается глутатионпероксидаза, восстановленный глутатион, витамин Е или другой антиоксидант [Birringer, EyTina, Salvatore, 2003], что ведет к прерыванию цепной реакции. Благодаря наличию в организме антирадикальной защиты, процессы пероксидации ограничиваются [Зенков, Ланкин, Меньщикова, 2004]. Физиологическая роль перекисного окисления заключается в участии в процессах самообновления, самоперестройки биологических мембран, ионного транспорта, регуляции активности мембраносвязанных ферментов и других физиологических реакций [Galeotti, Masotti, Borello, 2003]. При интенсивном воздействии на организм химических или физических факторов процессы ПОЛ многократно усиливаются, образование АФК возрастает, а когда происходит срыв механизмов антирадикальной защиты, развивается окислительный стресс, который может проявляться на клеточном, тканевом и организменном уровнях. При этом усиление перекисного окисления липидов, чрезмерная продукция органических перекисей приводят к развитию патологических процессов [Суханова, 2004; Барабой с соавт., 2004].
Повышение уровня ПОЛ наблюдается при многих заболеваниях, различных патологических состояниях и интоксикациях. При этом считают, что в случае болезни и интоксикации происходит возрастание ПОЛ, продукты которого и являются ответственными за повреждение клеток и тканей. Однако, как было установлено [Halliwell, Gutteridge, 2004], поврежденные ткани подвержены перекисному окислению липидов в гораздо большей степени, чем здоровые. Причина этого явления – инактивация некоторых биоантиоксидантов, утечка антиоксидантов из поврежденных клеток и выделение ионов металлов (особенно железа и меди) из мест их накопления в клетках и из металлопротеинов, гидролизованных освободившимися из разрушенных липосом ферментами [Воскресенский, Левицкий, 2003].
Состав продуктов перекисного окисления липидов достаточно сложен. К продуктам цепной реакции ПОЛ, прежде всего, относятся разновидности гидроперекисей, которые способны подвергаться нерадикальным окислительным превращениям, что приводит к образованию первичных (диеновые коьюгаты), и конечных продуктов ПОЛ (малоновый диальдегид, основания шиффа) [Журавлева с соавт., 2003].
Избыточное образование продуктов пероксидации приводит к целому комплексу цитотоксических эффектов, включающих инактивацию ряда ферментных систем, угнетение синтеза белка, повреждение и угнетение синтеза ДНК, лизис клеточных структур, нарушение репродуктивных процессов деления клеток.
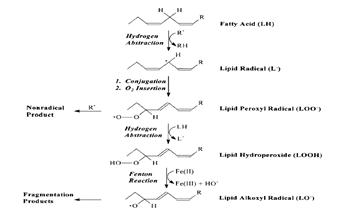
Рис.2. Радикальные и нерадикальные продукты ПОЛ [Aibert W, Girotty, 1998]
Особенно важно подчеркнуть, что происходит нарушение мембранных структур, изменение транспорта ионов Са2+, создающее угрозу жизни клетки [Michiel, Remacle, 2004].
Образование АФК, известных как прооксиданты, наблюдается во многих метаболических процессах и является обязательным атрибутом нормальной аэробной жизни. Функционирование и развитие клеток, а так же организма в целом, в кислородсодержащем окружении не могло бы быть возможным без существования защитных систем, основу которых составляют ферментативные и неферментативные антиоксиданты [Петрович, Гуткин, 2005]. Постоянное образование прооксидантов в живых организмах уравновешено их дезактивацией антиоксидантами, поэтому для поддержания гомеостаза необходима непрерывная регенерация антиоксидантной способности. Отсутствие или сбои этой непрерывности сопровождаются накоплением окислительных повреждений и приводят к возникновению окислительного стресса [Меньщикова с соавт., 2006].
Основными функциями антиоксидантной системы являются: ограничение интенсивности реакции свободнорадикального и перекисного окисления; защита чувствительных к окислительным повреждениям биомолекул мембран, внутри - и внеклеточных структур от действия свободных радикалов и перекисных соединений; восстановление окислительных молекулярных повреждений [Шепелев с соавт., 2004]. В целом основная задача системы антиоксидантной защиты состоит в предотвращении и ограничении развития патологических состояний, вызываемых окислительными повреждениями структур организма [Владимиров, 1998; Журавлев, 2003].
Общепринятой номенклатуры антиоксидантов в настоящее время нет. По химической природе биоантиокислители представляют собой широкий класс соединений: ферменты (СОД, каталаза, глутатионпероксидаза (ГПО)), фенолы и полифенолы (токоферолы, эвгенол), флавоноиды (рутин, кверцетин), стероидные гормоны и многие другие соединения [Бурлакова, Храпова, 2004]. В зависимости от растворимости различают жирорастворимые (витамин Е, А, К, убихинон) и водорастворимые (витамин С, SH-содержащие соединения) биоантиокислители, по молекулярной массе выделяют группу низкомолекулярных антиоксидантов (глутатион, α-токоферол, мочевая кислота) и высокомолекулярных ( ферритин, каталаза) [Кения, Лукаш, Гуськов, 1993].
К числу энзимных антиоксидантов относят прежде всего супероксидредуктазу (СОР), восстанавливающую О2- в пероксид водорода, СОД, катализирующую реакцию дисмутации О2- с образованием пероксида водорода и молекулярного кислорода [Cord, Fridovich, 2000], каталазу, восстанавливающую Н2О2, глутатионзависимые пероксидазы и трансферазы (ГТ) [Wendel, 2004].
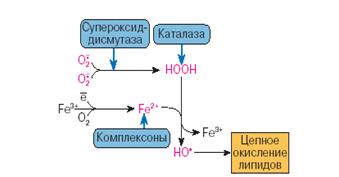
Рис.3. Антиоксиданты водной фазы [Владимиров, 1998]
Главной ферментативной системой плазмы крови является ГПО внеклеточных жидкостей и ГПО гидроперекисей липидов (ГПО 4), которая будучи липофильным соединением эффективно взаимодействует с гидроперекисями фосфотидилхолина, холестерина и эфиров холестерина в липопротеинах низкой плотности (ЛПНП), восстанавливая их, следовательно, защищая от окислительной модификации [Thomas at al., 2006; Abiaka, Al-Awadi, Olusi, 2003]. Кроме того ГПО 4 совместно с токоферолом практически полностью подавляет ПОЛ в биологических мембранах благодаря тому, что витамин Е эффективно восстанавливает пероксирадикалы, а фермент разлагает гидроперекиси, препятствуя тем самым их вовлечению в окислительный цикл [Ursini, Bindoli, 2005; Бондарь, Ланкин, Антоновский, 1989].
ГТ, в отличие от селенсодержащей ГПО, для которой лучшими субстратами являются гидрофильные гидроперекиси с малым размером молекул, эффективно восстанавливает гидрофобные гидроперекиси с большим объемом молекулы: гидроперекиси полиненасыщенных жирных кислот (линолевой и арахидоновой), фосфолипидов [Бобырев, Почерняева, Стародубцев, 2005; Колесниченко, Кулинский, 2004].
Вместе с тем во всех водных и липидных фазах организма могут протекать радикальные окислительные процессы, в защите от которых важную роль играют антиоксиданты-ингибиторы органических радикалов, среди которых важное место занимают соединения фенольного типа. В настоящее время выделено несколько тысяч фенольных соединений, среди которых выраженным антиоксидантным эффектом обладают витамины Е и К, убихиноны, триптофан и фенилаланин, а так же большинство растительных (флавоноиды) и животных пигментов [Дубина, 2005].
В антирадикальной защите липопротеинов плазмы крови и клеточных мембран α-токоферолу принадлежит ведущая роль – одна его молекула защищает ≈10000 молекул ненасыщенных жирных кислот [Евстигнеева, Волков, Чудинова, 2003], при этом считается, что α-токоферол способен обезвредить не менее 60% образующихся пероксильных радикалов [Тиунов, 1995]. Окисление α-токоферола со свободными радикалами компенсируется биорегенерацией молекул этого антиоксиданта в реакциях восстановления коантиоксидантами (АН), редокс-потенциал которых ниже, чем у радикала α-токоферола (α-Тф-О.) [Бурлакова, Крашков, Храпова, 1998]. В результате такой реакции не только происходит восстановление витамина Е, но и предотвращается возможность инициации α-токофероксильным радикалом окисления липидов:
α-Тф-О. + RH→ α-Тф-ОH + R. (1)
α-Тф-О. + АH→ α-Тф-ОH + А. (2)
В физиологических условиях вторая реакция обычно превалирует над первой, так как константа скорости реакции α-Тф-О. с НЖК не превышает 103М-1с-1, в то время как для реакции (1) константа скорости может достигать значений 104-107М-1с-1.К наиболее изученным коантиоксидантам относятся убихинол, аскорбиновая кислота (АК), билирубин [Менщикова, Зенков, 1993].
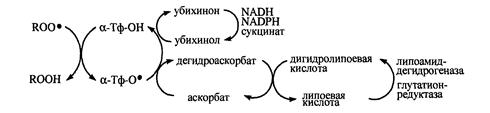
Рис.4. Окислительно-восстановителные превращения α-токоферола и сопряженных с ним коантиоксидантов [Меньщикова с соавт., 2006].
АК превосходит другие антиоксиданты плазмы в защите липидов от перекисного окисления, так как только это соединение достаточно реакционноспособно, чтобы эффективно ингибировать инициацию ПОЛ в водной фазе [Padayatty, Katz, Wang, 2003].
Важными внеклеточными антиокислителями являются альбумины – простые гидрофильные белки плазмы крови, которые являются основными носителями SН групп. SН содержащим соединениям принадлежит ведущая роль в защите клеток от ОН-радикала, образующегося в реакции Фентона или в результате разложения молекул воды под действием ионизирующих излучений.
Необходимо отметить наличие антирадикальных свойств у белков, хелатирующих ионы железа и других металлов с переменой валентностью. В первую очередь это трансферрин, лактоферрин и церулоплазмин. Основная роль трансферрина и близкого ему по структуре лактоферрина, содержащегося в молоке и выделяющегося фагоцитами при их активации, состоит в акцептировании «свободного» железа, что препятствует образованию радикалов НО.в реакциях Фентона, катализируемых ионами Fe [Зборовская, Банникова, 2000].Значимую роль в этом процессе играет церулоплазмин, Cu-содержащий белок, обеспечивающий окисление Fe 2+ до Fe3+ и делающий тем самым железо доступным для связывания трансферрина [Воскресенский с соавт., 2004; Krsek-Staples, Wbster, 2004; Василец, 2004]
Наиважнейшим антиоксидантом внеклеточной жидкости является мочевая кислота (МК). Ввиду высокого ее содержания в плазме крови человека некоторые исследователи считают, что на нее приходится 35-65% защиты липопротеинов от окисления, 10-15% ингибирования НО. и 12% ингибирования синглетного кислорода. Кроме того МК может выступать синергистом с радикалами α-токоферола и аскорбиновой кислотой, что усиливает их антиоксидантное действие [Булгакова, 2006].
Объектом исследования явилась замороженная плазма крови относительно здоровых небеременных и беременных женщин, поставляемая из Хакасского республиканского центра планирования семьи. Плазма хранилась при температуре -20° С до определения концентрации диеновых коньюгатов и малонового диальдегида. Всего было обследовано 127 женщин, деление на группы приведено в таблице 1.
Таблица 1
Группы обследованных женщин
| Сроки беременности | МДА, ДК |
| Контроль | N=31 |
| I триместр | N=29 |
| II триместр | N=34 |
| III триместр | N=33 |
2.2. Определение содержания малонового диальдегида.
В липидных системах в результате процессов ПОЛ образуется МДА, взаимодействие которого с 2-тиобарбитуровой кислотой (ТБК) приводит к образованию хромогена с максимумом поглощения в красной области видимого спектра при длине волны 532 нм. [Стальная, 1997].
Реактивы:
1. 30%-ная трихлоруксусная кислота (ТХУ);
2. 0,1 М этилендиаминтетрауксусная кислота (ЭДТА);
3. 1%-ная тиобарбитуровая кислота;
4. 0,05 н раствор NaOH;
5. Физиологический раствор.
Ход определения:
К 0,2 мл образца добавляли 0,8 мл физиологического раствора и 0,5 мл ТХУ. Перемешивали, оставляли стоять на льду 2 часа. Затем центрифугировали 15 мин. при 3000 об/мин. 1 мл супернатанта переносили в другую пробирку, добавляли 0,075 мл 0,1 М ЭДТА и 1%-ного ТБК, растворенной в 0,05 Н растворе NaOH. Содержимое перемешивали и ставили в кипящую водяную баню на 15 минут. Затем пробирки охлаждали при комнатной температуре и измеряли поглощение при длинах волн 532нм, а затем при 600 нм.
Расчет МДА производят по формуле:
D532 – D600
C= _________________ x f,
E
где:
C – концентрация МДА, мкмоль/л плазмы (эритроцитов),
D532 оптическая плотность при длине волны 532 нм;
D600 – оптическая плотность при длине волны 600 нм;
E – коэффициент экстинции 1,56x105М-1см-1;
f – коэффициент разведения. (f=9,94)
2.3. Определение содержания диеновых коньюгатов.
Вследствие π-π переходов спектры конъюгированных гидроперекисей полиненасыщенных жирных кислот характеризуются интенсивным поглощением в ультрафиолетовой области спектра с максимумом при 232-234 нм.
Определение содержания ДК проводили в экстрактах эритроцитов. Для этого липиды из эритроцитов экстрагировали стократным избытком смеси растворителей (гептан-изопропанольная смесь в соотношении 1:1). В гомогенизатор вносили 0,1 мл. упакованных эритроцитов, добавляли 5 мл. изопропилового спирта и тщательно растирали до получения гомогенной суспензии. Содержимое гомогенизатора количественно переносили в мерную центрифужную пробирку, в которую затем добавляли 5 мл. гептана.
Экстракт центрифугировали в течение 10 минут при 1700g. Надосадочную фракцию переносили в градуированную пробирку и добавляли 1/5 объема 0,74%-ного водного раствора KCI для отмывки липидного экстракта от нелипидных примесей. После тщательного встряхивания образовавшаяся эмульсия расслаивалась на две прозрачные фазы. В гептановом экстракте (верхняя фаза) измеряли спектрофотометрически содержание сопряженных диенов в кювете с длиной оптического пути 1,0 см против гептана. Расчет количества ДК производили, используя молярный коэффициент экстинкции при длине волны 233 нм, равный 27000 М¯¹*см¯¹ [Паранич и соавт., 1993] и выражали в ммолях на мл клеток.
С=Д/К*F
Д – оптическая плотность;
К – коэффициент ммолярной экстинкции = 21,0 мМ-1 см-1;
F – фактор разведения.
2.4. Статистическая обработка результатов
Статистическую обработку результатов осуществляли общепринятыми методами с использованием пакета прикладных программ Statistica 7.0. Достоверность различий оценивали по непараметрическому критерию Манна-Уитни, с достоверностью Р<0,05 [Лакин, 1980].
Глава 3. Результаты исследований и обсуждение.
Говоря об общей опасности воздействия свободных радикалов на организм, нельзя обойти эту проблему и у беременных женщин, от здоровья которых зависит будущее целого поколении детей.
При неосложненной беременности в организме женщин происходит целый ряд адапционно-приспособительных процессов, направленных на обеспечение адекватного течения гестационного периода, роста и развития плода. Известно, что важными составляющими адаптивных перестроек являются показатели агрегатного состояния крови, иммунного статуса, эндокринной системы а так же перекисного окисления липидов. В их сбалансированном взаимодействии в ответ на регулирующее влияние гипоталамуса заложено качество адаптации организма [Гусак, 2006].
Физиологическая беременность может сопровождаться существенными изменениями в про/антиоксидантном статусе. Одними из параметров, которые позволяют оценить состояние свободнорадикальных процессов является диеновые коньюгаты и малоновый диальдигид. В нормальных условиях активность этих процессов находится на невысоком уровне, но при стрессовых ситуациях происходит усиленная активация процессов ПОЛ под действием АФК, что приводит к патологическому состоянию.
3.1. Содержание продуктов перекисного окисления липидов в плазме крови небеременных женщин.
В данной работе было определено содержание диеновых коньюгатов и малонового диальдегида у небеременных женщин (N=31), которые составили группу контроля. Согласно литературным данным содержание продуктов ПОЛ в частности МДА, являющегося одним из конечных продуктов данного процесса, с возрастом неуклонно возрастает [Банкова, Никанорова, 1988]. В группе небеременных женщин возраст значительно варьировал: от 18 до 39 лет. В связи с этим, мы разделили женщин контрольной группы по возрастному критерию на две подгруппы: в первую подгруппу вошли женщины в возрасте 18-28 лет (N=18), во вторую – 29-39 лет (N=13).
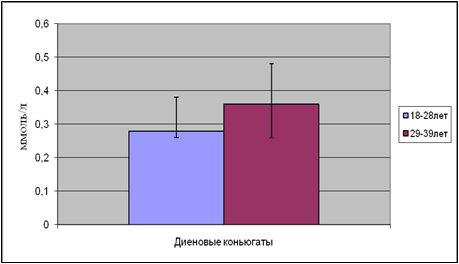 |
Содержание ДК и МДА в этих подгруппах приведены на рисунках 5 и 6.
Рис.5. Содержание диеновых коньюгатов в возрастных подгруппах
В возрастной подгруппе от 18 до 28 лет содержание ДК составило 0,28 ммоль/л, а в возрастной подгруппе от 29 до 39 лет концентрация увеличилась на 28%. Та же закономерность выявлена и в содержании МДА.
Концентрация МДА в первой подгруппе составила 0,85мкмоль/л, во второй подгруппе уровень МДА повысился на 31%.
Наблюдаемые изменения свидетельствуют о том, что с возрастом на фоне усиления процессов ПОЛ происходит угнетение антиоксидантной системы. Нарушения в системе перекисного окисления липидов и антиоксидантной активности являются одним из механизмов формирования антиоксидантной недостаточности вследствие чрезмерного усиления ПОЛ.
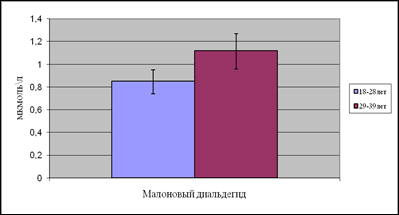
Рис.6. Содержание малонового диальдегида в возрастных подгруппах
В результате активации ПОЛ и накопления свободных радикалов происходит окислительная модификация липопротеинов плазмы крови, нарушение структурно-функциональной целостности клеточных мембран, освобождение лизосомальных ферментов, что в конечном итоге приводит к патологическим процессам в клетке и организме в целом.
При анализе содержания ДК и МДА между двумя возрастными подгруппами достоверных отличий не выявлено. Вследствие чего при сравнительном анализе содержания продуктов ПОЛ у беременных и небеременных женщин будут использованы данные по общей контрольной группе. Результаты исследования содержания ДК и МДА в общей контрольной группе приведены на рисунке 7.
Уровень ДК в контрольной группе женщин составил 0,35 ммоль/л, а МДА – 0,96 мкмоль/л. Наши данные по содержанию МДА в плазме крови согласуются с приведенными в литературе [Гусак, 2006]. Данных по содержанию ДК в плазме крови у относительно здоровых женщин мы в доступной литературе не встретили.
Анализ полученных результатов показал, что уровень ДК, являющихся первичными продуктами ПОЛ, существенно превышает таковой для конечного продукта – МДА.
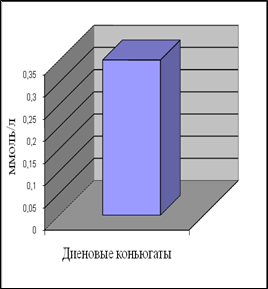
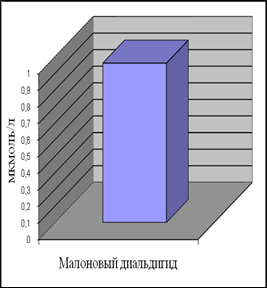
Рис.7. Содержание продуктов перекисного окисления липидов в плазме крови небеременных женщин
Согласно данным литературы, у женщин показатели перекисного окисления липидов, а так же антиоксидантной системы изменяются в динамике репродуктивного цикла (1, 7, 14, 21 дни) [Гусак, 2006]. В первые дни менструального цикла происходит увеличение соотношения адреналин/серотонин, что свидетельствует о преобладании в функциональной активности гипоталамуса адренергических компонентов. Это приводит к депрессии противосвертывающей и антиокислительной систем крови, а так же активации клеточного и гуморального иммунитета, повышению неспецифической иммунной резистентности. Что проявляется увеличением концентрации диеновых коньюгатов и малонового диальдегида, активацией свертывающего потенциала крови.
К 21 дню цикла активность АОС прогрессивно увеличивается, на фоне этого снижается количество продуктов ПОЛ, в том числе ДК и МДА. Это необходимо для обеспечения процессов имплантации в слизистую матки, развития плаценты и создания оптимальных условий, которые позволят, с одной стороны – обеспечить надежную защиту организма женщины, а с другой – вынашивание беременности. Данная закономерность имеет глубокий биологический смысл и направлена на обеспечение условий для реализации репродуктивной функции человека.
В данной работе мы не учитывали возможность влияния динамики репродуктивного цикла женщин на уровень продуктов ПОЛ, хотя это явление могло отразиться на полученных результатах.
Во время гестации, начиная с момента зачатия и до завершения родов, в организме матери возникают интегративные процессы, которые необходимы для поддержания функционального единства организма матери и плода. Эти процессы позволяют выполнить главную задачу, то есть сохранение плода, и заключается в адаптации в I триместре беременности.
Несмотря на то, что в антигенном отношении мать и плод всегда несовместимы, в большинстве случаев после имплантации бластоцисты беременность развивается нормально и завершается родами в срок. Если бы взаимоотношения между матерью и плодом строились по варианту реципиент – аллотрансплантат, то беременность вряд ли продолжалась дольше срока, чем выживание обычного трансплантата. Следовательно, в системе "мать – плод" существуют механизмы, направленные на сопереживание двух антагонистически настроенных субъектов.
В данной работе было определено содержание ДК и МДА в плазме крови у беременных женщин (N=96), разделенных по триместрам, в возрасте от 20 до 28 лет.
Группу сравнения составили небеременные женщины. Результаты определения уровня ДК и МДА приведены на рисунках 8 и 9.
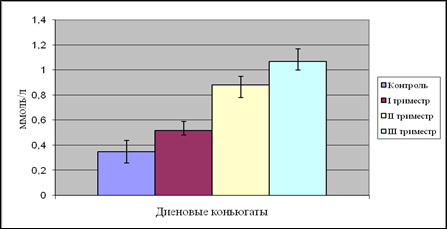 |
Рис. 8. Содержание диеновых коньюгатов в плазме крови женщин в динамике беременности
У женщин в I триместре беременности наблюдается достоверное повышение уровня продуктов ПОЛ по сравнению с контролем. Так, содержание ДК и МДА повысилось на 74 и 77% соответственно. Это можно объяснить тем, что сразу же после зачатия происходит значительная перестройка жизнедеятельности организма беременных женщин, которая сопряжена с изменениями в системах крови, гемостаза, эндокринной, иммунной системах и с изменением биохимического состояния организма в целом. «Не ожидая» подобных изменений, ткани и органы испытывают определенный стресс, в результате которого резко повышается количество свободных радикалов, атакующих, помимо прочего, клетки плаценты и эмбриона.
Увеличение количества свободных радикалов (СР) во время беременности связано с различными причинами. Одной из причин является их участие в синтезе прогестерона. СР активируют процесс перекисного окисления липидов, в результате которого образуются гидроперекиси холестерина, являющиеся предшественниками данного гормона.
Прогестерон является очень важным гормоном, необходимым для поддержания всего течения беременности. Он подготавливает эндометрий матки к имплантации оплодотворенной яйцеклетки, а затем способствует сохранению беременности: подавляет активность гладкой мускулатуры матки, поддерживает в центральной нервной системе доминанту беременности; стимулирует развитие концевых секреторных отделов молочных желез и рост матки, синтез стероидных гормонов; оказывает иммунодепрессивное действие, подавляя реакцию отторжения плодного яйца [Савченко с соавт., 2006].
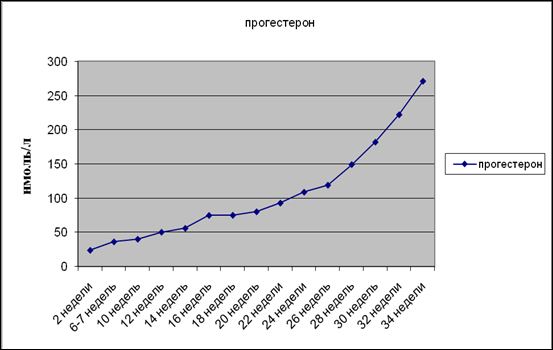
Рис 9. Концентрация прогестерона при разных сроках беременности
Кроме того, прогестерон является предшественником стероидных гормонов плода, а так же эстрогенов, андрогенов, альдостерона и других гормонов коры надпочечников. Содержание прогестерона в крови беременной женщины увеличивается, повышаясь в 2 раза к 7-8 неделе, а затем более плавно возрастает к 34 недели (Рис 9). Следовательно, интенсификация процессов перекисного окисления липидов во время беременности является физиологически необходимым процессом.
Гормональные связи «пронизывают» все компоненты функциональной системы мать-плацента-плод. Так, в I триместре беременности происходит тесное взаимодействие материнско-плацентарной эндокринной системы, а во II и особенно в III триместрах плод и плацента выступают как общий орган синтеза эстрогенов.
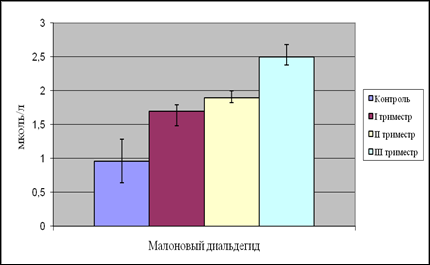
Рис. 9. Содержание малонового диальдегида в плазме крови женщин в динамике беременности
Дальнейший анализ полученных нами данных показал, что уровень как МДА, так и ДК во II триместре беременности продолжает достоверно расти как по сравнению с группой контроля, так и c I триместром беременности.
Концентрация ДК во II триместре беременности увеличилась по сравнению с группой небеременных женщин и I триместром на 151 и 69% соответственно. Содержание МДА возросло на 97 и 11% так же по сравнению с контролем и I триместром беременности.
Сравнение уровней содержания первичных и вторичных продуктов ПОЛ во II триместре беременности по сравнению с I триместром показал более выраженный прирост концентрации диеновых коньюгатов по сравнению с малоновым диальдегидом. Этому явлению можно найти несколько объяснений.
Во-первых, из литературных данных известно, что МДА может метаболизироваться. В этом процессе принимает участие альдегиддегидрогеназа. Наличие такого фермента отмечено в плазме крови [Соловьева, 2007].
Во-вторых, МДА является чрезвычайно реакционноспособным соединением, способным вступать в образование шиффовых оснований с соединениями, содержащими NH2-группы. К такому роду соединений относятся аминокислоты, низкомолекулярные пептиды и прочие соединения. В этом случае часть МДА извлекается из пула свободного МДА, образуя так называемый «связанный» МДА, концентрация которого, как показали некоторые исследования, повышается при ряде патологических состояниях. Используемая нами методика позволяет определить только свободный МДА без учета связанного МДА.
В-третьих, нельзя исключить, что метаболизм продуктов липопероксидации во II триместре такой, что скорость превращения ДК в МДА снижена.
В III триместре содержание ДК в плазме крови беременных женщин достигает своего максимума и составляет 1,07 ммоль/л. Уровень МДА так же повышается по сравнению с контролем и вторым триместром на 160 и 31% соответственно.
Согласно литературным данным повышение продуктов ПОЛ в III триместре беременных обусловлено угнетением ферментативных и неферментативных механизмов антиперекисной защиты. Происходит снижение активности каталазы [Гусак, 2007] и церулоплазмина (ЦП) [Качалина, Морозова, 2003]. ЦП обладает ферроксидазной активностью, а так же ингибирует супероксидный анион-радикал, избыток которого приводит к усилению процесса ПОЛ.
Кроме того, в плазме крови церулоплазмин совместно с трансферрином образует антиоксидантную систему, регулирующую концентрацию восстановленных ионов железа, и суммарная антиокислительная активность плазмы крови в отношении Fe2+ -индуцированного ПОЛ в основном определяется содержанием в ней данных компонентов [Меньщикова, 2006].
Так же в III триместре беременности происходит возбуждение адренергических структур гипоталамуса, что приводит к увеличению соотношения адреналин/серотонин, депрессии антиоксидантного потенциала плазмы крови и активации иммуногенеза [Гусак, 2007].
Усиление процессов перекисного окисления липидов в крови беременных женщин может быть связано с различными причинами.
Чрезмерное и бесконтрольное увлечение препаратами железа может стать причиной увеличения числа свободных радикалов, так как любой антиоксидант в определенных условиях может выступать прооксидантом, инициируя окислительные процессы [Меньщикова с соавт., 2006]. В то же время дефицит железа может способствовать развитию железодефицитной анемии (ЖДА), следствием которой является тканевая гипоксия [Мусаев, 2004].
По данным литературы длительная гипоксия приводит к истощению АОС, что влечет за собой неконтролируемый процесс липопероксидации и накопление продуктов свободно-радикального окисления, оказывающих неблагоприятное влияние на систему мать-плацента-плод [Евсюкова, Савельева, 2005].
Другой причиной является образование активных форм кислорода при применении лекарственных средств. Подвергаясь всевозможным ферментативным превращениям в организме, молекулы некоторых лекарств теряют свои электроны в этих химических реакциях, превращаясь в свободные радикалы.
Состояние стресса так же оказывает мощное влияние на активацию процессов ПОЛ. Гормоны стресса, адреналин и кортизол, при неблагоприятных жизненных ситуациях вырабатываются в повышенных количествах, нарушая питание и нормальное дыхание клетки, что моментально приводит к накоплению и распространению радикалов во всем организме.
Так же важно отметить, что беременные женщины подвергаются регулярному влиянию неблагоприятных факторов окружающей среды: загрязненный воздух, табачный дым, ультрафиолетовое излучение. Это усугубляет процесс физиологического течения беременности при условии дефицита антиокислителей. Кроме того данные литературы свидетельствуют о корреляции окислительного напряжения организма матери и неблагоприятным результатом беременности, в том числе выкидыше[Young-Ju Kim, Yun-Chul Hong, Kwan-Hee Lee, 2005].
1. Содержание диеновых коньюгатов в плазме крови небеременных женщин составило 0,26-0,54 ммоль/л.
2. Содержание малонового диальдегида в плазме крови небеременных женщин составило 0,64-1,28 мкмоль/л.
3. Уровень ДК в плазме крови в I и II триместрах беременности достоверно повышается на 74 и 151% соответственно по сравнению с группой контроля. В III триместре концентрация диеновых коньюгатов достигает своего максимума и составляет 1,07 ммоль/л.
4. Содержание МДА в плазме крови в I, II, III триместрах беременности прогрессивно увеличивается на 77, 97, 160% соответственно по сравнению с группой небеременных женщин.
Банкова В.В. Деградация малонового диальдегида в эритроцитах и ее возрастные, сезонные и суточные изменения / В.В. Банкова, Т. М. Никанорова // Вопр. мед. Химии, 1988. - № 6. - С. 27-29.
Барабой В.А. Механизмы стресса и перекисное окисление липидов / В.А. Барабой // Успехи соврем. Биологии, 1991. - Т. 111. - № 6. - С. 923-931.
Барабой В.А. Перекисное окисление и стресс / В.А. Барабой, И.И. Брехман, В.Г. Голожин с соавт. - М.: Наука, 2004. - 148 с.
Бобырев В.Н. Специфичность систем антиоксидантной защиты органов и тканей – основа дифференцированной фармакотерапии антиоксидантами / В.Н. Бобырев, В.Ф. Почерняева, С.Г. Стародубцев // Экспериментальная и клиническая фармакология, 2005. – Т. 57. - №1. - С. 78-86.
Бондарь Т.Н. Восстановление органических гидроперекисей глутатионпероксидазой и глутатион-S-трансферазой: влияние структуры субстрата / Т.Н. Бондарь, В.З. Ланкин, В.А. Антоновский // Докл. АН СССР, 1989. - T. 304. - №1. – С. 217-220.
Булгакова Е.Б. Перекисное окисление липидов мембран и природные антиоксиданты / Булгакова Е.Б. // Успехи химии, 2006. - № 9. – 250 c.
Бурлакова Е.Б. Перекисное окисление липидов мембран / Е.Б. Бурлакова, Н.Г. Храпова // Успехи химии, 2004. - Т. 54. - C. 1540-1558.
Бурлакова Е.Б. Роль токоферолов в пероксидном окислении липидов биомембран / Е.Б. Бурлакова, С.А. Крашков, Н.Г. Храпова // Биологические мембраны, 1998. - Т. 15, № 2. - С. 137-167.
Василец И.М. Церулоплазимины. Их молекулярная структура и биологические функции / И.М. Василец // Успехи биол. Химии, 2004. - № 14. - С. 172-200.
Владимиров Ю.А. Перекисное окисление липидов в биомембранах / Ю.А. Владимиров, А.И. Арчаков. - М.: Наука, 2003. – С. 230-272.
Владимиров Ю.А. Свободные радикалы в живых системах / Ю.А. Владимиров, О.А. Азизова, А.И. Деев с соавт. // Итоги науки и техники, 2000. – Т. 29. - С. 151-167.
Владимиров Ю.А. Свободные радикалы и антиоксиданты / Ю.А. Владимиров // Вестн. РАМН, 1998. - № 7. - С. 43-51.
Воейков В.Л. Благотворная роль активных форм кислорода / В.Л. Воейков // Биохимия, 2004. - № 1 - С. 27-38.
Воскресенский О.Н. Перекиси липидов в живом организме / О.Н. Воскресенский, А.П. Левицкий // Вопр. мед. Химии, 2003. - Т. 16. - № 6. – С. 563-583.
Воскресенский С.К. Антиоксидантная система, онтогенез и старение / С.К. Воскресенский, И.А. Жутаев, В.Н. Бобырев с соавт. // Вопр. мед. Химии, 2004. - № 1. - C. 14-27.
Гусак Ю.К. Психонейроиммунологические особенности адаптивных механизмов нормального репродуктивного цикла у женщин [Электронный ресурс] / Ю.К. Гусак, Ю.В. Лазарева, В.Н. Морозов, 2006. – Режим доступа: http:www.mednet.сom
Гусак Ю.К. Психонейроиммунологические особенности адаптивных механизмов при нормально протекающей беременности [Электронный ресурс] / Ю.К. Гусак, Ю.В. Лазарева, В.Н. Морозов, 2007. - Режим доступа: http:www.mednet.сom
Дубина Е.Е. Некоторые особенности функционирования ферментной антиоксидантной защиты плазмы крови человека / Е.Е. Дубина // Биохимия, 2005. - Вып. 2. – С. 3-18.
Дубинина Е.Е. Биологическая роль супероксидного анион-радикала и супероксиддисмутазы в тканях организма / Е.Е. Дубинина // Успехи современной биологии, 2004. - Т. 108. - №1. – C. 3-17.
Евстигнеева Р.П. Витамин Е как универсальный антиоксидант и стабилизатор биологических мембран / Р.П. Евстигнеева, И.М. Волков, В.В. Чудинова // Биол. Мембраны, 2003. - № 2. С. 119-137.
Евсюкова И.И. Свободнорадикальное окисление у доношенных новорожденных детей с различной патологией / И.И. Евсюкова, Т.В. Савельева // Педиатрия, 2005. - №1. – С. 13-16.
Журавлев А.И. Биоантиокислители в живом организме / А.И. Журавлев. - М.: Наука, 2003. - C. 19-30.
Журавлева Т.Д. Возрастные особенности свободнорадикального окисления липидов и антиоксидантной защиты в эритрацитах здоровых людей / Т.Д. Журавлева, С.Н. Суплотов, Н.С. Киянюк с соавт. // Вопр. мед химии, 2003. - № 5. - C. 17-18.
Зборовская В.А. Антиоксидантная система организма, ее значение в метаболизме / В.А. Зборовская, М.В. Банникова // Вестник РАМН, 2000. - № 6. - С. 53-63.
Зенков Н.К. Окислительный стресс / Н.К. Зенков, В.З. Ланкин, Е.Б. Меньщикова. - М.: Наука, 2004. - 343с.
Качалина Т.С. Прогностическая значимость определения церулоплазмина в третьем триместре беременности [Электронный ресурс] / Т.С. Качалина, Т.А. Морозова, 2006. – Режим доступа: http: www.iprit.ru/ chemical agents action=1179
Кения М.П. Роль низкомолекулярных антиоксидантов при окислительном стрессе / М.П. Кения, А.И. Лукаш, Е.П. Гуськов // Успехи современной биологии, 1993. - Т. 113. - № 4. - С. 456-468.
Колесниченко Л.С. Глутатионтрансферазы / Л.С. Колесниченко, В.И. Кулинский // Успехи совр. Биологии, 2004. - Т. 107. - вып. 2. - С. 179-193.
Лакин Г.Ф. Биометрия / Г.Ф. Лакин. - М.: Высш. школа, 1998. - 293с.
Меньщикова Е.Б. Антиоксиданты и ингибиторы радикальных окислительных процессов / Е.Б. Меньщикова, Н.К. Зенков // Успехи соврем. Биологии, 1993. - Т. 113. - №4. - C. 442-453.
Меньщикова Е.Б. Окислительный стресс. Прооксиданты и антиоксиданты. / Е.Б. Меньщикова, В.З. Ланкин, Н.К. Зенков с соавт. - М.: «Слово», 2006. – 553 с.
Мусаев А.Т. Диагностика гипоксии плода по данным показателей перекисного окисления липидов и антиокислительной активности / А.Т. Мусаев // Педиатрия, 2004. - № 12. – С. 88-96.
Осипов А.Н. Активные формы кислорода и их роль в организме / А.Н. Осипов, О.А. Азизова, Ю.А. Владимиров // Успехи соврем. биологии, 2003. - Т. 31. - C. 180-208.
Осипов А.Н. Образование гидроксильных радикалов при взаимодействии гипохлорита с ионами железа / А.Н. Осипов, Э.Ш. Якутова, Ю.А. Владимиров // Биофизика, 2003. – Т. 38. - вып 3. – С. 390-396.
Петрович Ю.А. Свободнорадикальное окисление и его роль в патогенезе воспаления, ишемии и стресса / Ю.А. Петрович, Д.В. Гуткин // Патол. физиол. и экперим. Терапия, 2005. - № 5. - С. 85-92.
Савченко А.А. Система мать-плацента-плод. Вопросы функциональной диагностики / А.А. Савченко, Н.М. Титова, Л.А. Новикова с соавт. - Красноярск: РИО КрасГУ, 2006. – 129 с
Соловьева А.Г. Активность альдегиддегидрогеназы в эритроцитах, тромбоцитах и плазме крови крыс в норме и при ожоге / А.Г. Соловьева // Успехи соврем. Естествознания, 2007. - № 12. – С. 12-15.
Стальная И.Д. Метод определения диеновой коньюгации ненасыщенных высших жирных кислот / И.Д. Стальная // Современные методы в биохимии под ред. Ореховича В.Н., 1997. - С. 63-64.
Стальная И.Д. Метод определения малонового диальдегида / И.Д. Стальная, Т.Г. Гаришвили // Современные методы в биохимии под ред. Ореховича В.Н., 1997. - C. 66-68.
Суханова Т.А Патохимия клетки / Т.А. Суханова // Успехи соврем. биологии, 2004. – Т. 40. – С. 82-104.
Тиунов Л.А. Механизмы естественной детоксикации и антиоксидантной защиты / Л.А. Тиунов // Вестн. РАМН, 1995. - № 3. - C. 9-13.
Хавинсон В.Х. Свободнорадикальное окисление и старение / В.Х. Хавинсон, В.А. Баринов, А.В. Арутюнян с соавт. - СПб.: Наука, 2003. - С. 10-122.
Шепелев А.П. Роль процессов свободнорадикального окисления в патогенезе инфекционных болезней / А.П. Шепелев, И.В. Корниенко, А.В. Шестопалов с соавт. // Вопр. мед. Химии, 2004. - № 2. - С. 15-17.
Якутова Э.Ш. Образование свободных радикалов при взаимодействии гипохлорита с ионами железа / Э.Ш. Якутова, Е.С. Дремина, С.А. Евгина с соавт. // Биофизика, 2004. – Т. 39. - вып. 2. – С. 275-279.
Abiaka C. Effect of Prolonged Storage on the Activities of Superoxide Dismutase, Glutathione Reductase, and Glutathione Peroxidase / C. Abiaka, F. Al-Awadi, S. Olusi // Clinical Chemistry, 2003. - Vol. 46. - Issue 4 - P. 560-576.
Auchere F. What is the ultimate fate of superoxide anion in vivo? / F. Auchere, F. Rusnak // J. Biol. Inord. Chem., 2005. - Vol. 7 - P. 664-667.
Beyer R. E. The participation of coenzim Q in free radical production and antioxidation / R. E Beyer // Free Radic. Biol. Med., 2004. – Vol. 8. – P. 545-565.
Birringer M. Vitamin E analogues as inducers of apoptosis: structure-function relation / M. Birringer, J. H. EyTina, B A. Salvatore // Br. J. Cancer., 2003. – Vol. 88. – P. 1948-1955.
Carole B. Rudra A prospective study of early-pregnancy plasma malondialdehyde concentration and risk of preeclampsia / Carole B. Rudra, Chunfang Qiu, Robert M. David at аl. // Clinical Biochemistry, 2006. Vol. 39. – P. 722-726.
Chen R. Microdialysis sampling combined with electron spin resonance for superoxide radical detection in microliter samples / R. Chen, J. T. Warden, J. A. Stonken // Anal. Chem., 2004. - Vol. 76. - P. 4734-4740.
Cord J.M. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein) / J. M. Cord, I. Fridovich // J. Biol. Chem., 2000. - Vol. 244. - Issue 22. - P. 6049-6055.
Dix T.A. Mechanisms and biological significance of lipid peroxidation initiation / T.A. Dix, J. Aikens // Chem. Res. Toxicol., 2005. - Vol. 6. - Р. 2-18.
Eisenberg W. C. Cytogenetic effects of singlet oxygen / W. C. Eisenberg, K. Taylor, R. R. Guerrero // J Photochem. Photobiol., 2002. – Vol. 16. – P. 381-384.
Galeotti T. Oxy-radical metabolism and control of tumour growth / T. Galeotti, L. Masotti, S. Borello // Xenobiotika, 2003. – Vol. 21. – P. 1041-1052.
Gutteridge J. M. C. Lipid peroxidation and antioxidation as biomarkers of tissues damage / J. M. C. Gutteridge // Clinikal Chemistry, 2005. - Vol. 41. - №12. - P. 1819-1828.
Halliwell B. Oxygen toxiciti, oxygen radicals, transition metals and disease / B. Halliwell, J. M. C Gutteridge // Biochem., 2004. – Vol .215. - P. 1-14.
Kira, Y. Association of Cu-Zn-type superoxide dismutase with mitochondria and peroxisomes / Y.Kira, E. F. Sato, M. Inoue // Arsh. Biochem. Biophys., 2003. – Vol. 399. – P. 96-102.
Klebanoff, S. J.Myeloperoxidase: role in neutrophil – mediated toxicity / S. J. Klebanoff // Molecular Biologi and Infectious Diseases., 2006. – Vol. 24. - P. 283-289.
Krsek-Staples, J.A. Ceruloplasmin inhibits carbonyl formation in endogenous cell proteins / J. A. Krsek-Staples, R. O. Wbster // Free Radic Biol. Med., 2004. - Vol. 14. - Р. 115-25.
Maiorino, M. Prooxidant role of vitamin E in copper induced lipid peroxidation / M. Maiorino, A. Zamburlini, A. Roveri // FEBS Lett., 2005. – Vol. 330. – P. 174-176.
Melov, S. Animal models of oxidative stress, aging and therapeutic antioxidant interventions / S. Melov // Int. J. Biochem. Cell Biol., 2003. – Vol. 34. – P. 1395-1400.
Meral, A. Lipid peroxidation and antioxidant status in beta-thalassemia / A. Meral, P. Tuncel, E. Surmen-Gur // Pediatr. Hematol. Oncol., 2000. - Vol. 17. - P. 687-693.
Michiels, C. Cytotoxicity of linoleic acid peroxide, malondialdehyde and 4-hydroxynonenal towards human fibroblast / C. Michiels, J. Remacle // Toxicology, 2004. - Vol. 66. - №2. - P. 225-234.
Okado-Matsumoto, A. Subcellular distribution of superoxide dismutases in rat liver: Cu,Zn-SOD in mitochondria / A. Okado-Matsumoto, I. Fridovich // J. Biol. Chem., 2003. - Vol. 276. - P. 38388-38393.
Padayatty, S. J. Vitamin C as an Antioxidant: Evaluation of Its Role in Disease Prevention / S. J. Padayatty, A. Katz, Y. Wang // Journal of the American College of Nutrition, 2003. - Vol. 22. - Р. 18-35.
Thomas, J. P. Enzymatic reduction of phospholipid and cholesterol hydroperoxides in artificial bilayers and lipoproteins / J. P. Thomas, P. G. Geiger, M. Maiorino et al. // Biochim. Biophys. Acta., 2006. – Vol. 1045. – P. 252-260.
Tohoku, J. Nitric Oxide, Lipid Peroxides, and Uric Acid Levels in Pre-Eclampsia and Eclampsia / J Tohoku // Biochem., 2004. – Vol. 15. - P. 87-92.
Ursini, F. The role of selenium peroxidases in the protection against oxidative damage of membranes / F. Ursini, A. Bindoli // Chem. Phys. Lipids., 2005. – Vol. 44. – P. 255-276.
Wendel, T. Enzimes acting against reactive oxygen / A Wendel // Enzymes, 2004. – Vol. 34. – P. 161-167.
Young-Ju Kim Oxidative stress in pregnant women and birth weight reduction / Young-Ju Kim, Yun-Chul Hong, Kwan-Hee Lee // Reproductive Toxicologi, 2005. – Vol. 19. – P. 487-492.
The present work is devoted to the mechanisms of lipid peroxidation and antioxidant defence. A special attention is paid to the experimental research of the products of lipid peroxidation.
The purpose of this work is measuring of the level of MDA and DС in blood plasma of women in different stages of pregnancy.
The maintenance of a recreation center DС and MDA in plasma of blood of not pregnant women has made 0,26 - 0,54 mmol/l and 0,64-1,28 µmol/l accordingly.
The level of a recreation center in plasma of blood in I and II trimesters of pregnancy authentically raises on 74 and 151 % accordingly in comparison with group of the control. In III trimester concentration DС reaches the maximum and makes 1,07 mmol/l.
Maintenance MDA in plasma of blood in I, II, III trimesters of pregnancy progressively increases for 77, 97, 160 % accordingly in comparison with group of not pregnant women.
© 2009 База Рефератов