
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Реферат: Противостояние организма изменениям барических и термических условий
Реферат: Противостояние организма изменениям барических и термических условий
Реферат
по биологии
на тему:
«Противостояние организма изменениям барических и термических условий»
Приспособление к гипоксии
Мы уже знаем, что экстренное приспособление к высотной гипоксии вызывает прежде всего ряд физиологических реакций, направленных на возможно более полное удовлетворение потребности организма в кислороде. Но этого недостаточно даже на сравнительно небольших высотах: уже на уровне 3500 м парциальное давление кислорода в альвеолах легких снижается на 53%, а насыщение им крови – на 20%.
Разные органы и составляющие их клетки в неодинаковой степени чувствительны к недостатку кислорода. Особенно страдает от этого центральная нервная система, в частности нервные клетки ее высшего отдела – коры головного мозга. На небольших высотах человек этого еще не замечает, но инструментальные физиологические исследования показывают, что уже на высоте 1500 м у человека ухудшается темновая адаптация, т.е. приспособление глаза к ясному зрению при переходе от яркого освещения к полутемноте. На высотах 2000–2200 м отмечаются нарушения и в функциях коры головного мозга: увеличивается скрытый период речевых реакций, а сама речь становится более примитивной, нарушается тонкая дифференцировка восприятий. На высоте 4000 м хуже протекают условные рефлексы, а выше 6000 м в коре мозга возникает разлитое торможение, появляется сонливость и, наконец, теряется сознание. Но ведь на высоте надо и двигаться, и нередко совершать значительную работу. А это еще более увеличивает потребность в кислороде.
Общеизвестно, что энергию, необходимую для всех проявлений жизнедеятельности, организм получает путем окисления различных веществ, прежде всего углеводов и жиров, накапливая ее посредством окислительного фосфоридирования в легкоутилизируемой форме АТФ. Процесс окисления заключается в отнятии водорода от окисляемых веществ и перенесении его протона и электрона через дыхательную цепь на кислород. Перенос каждой пары протонов и электронов сопровождается образованием трех молекул АТФ. При этом, чем богаче водородом окисляемое вещество, тем больше выход АТФ: при окислении глюкозы образуется 38 молекул АТФ, а при окислении жирных кислот, например пальмитиновой, – 136. А если кислорода недостаточно? Если протоны и электроны движутся по дыхательной цепи, а отдавать их некому? Если вся дыхательная цепь оказывается восстановленной, т.е. забита электронами и протонами? Тогда на помощь «спешит» гликолиз.
Гликолиз происходит следующим образом. Глюкоза подвергается фосфорилированию. Образовавшийся глю-козо-фосфорный эфир после ряда промежуточных превращений и внутримолекулярных перегруппировок расщепляется на две молекулы фосфотриозы, которые подвергаются окислению. От них отнимается водород, но принимает его не кислород, а кофермент НАД. Конечным же продуктом является ПВК. Дальнейшая судьба этого соединения неодинакова и зависит от степени обеспечения организма кислородом: если его достаточно, то ПВК окисляется до СО2 и H2O, а водород с НАД · H2 через дыхательную цепь переносится на кислород; при нехватке кислорода НАД · H2 отдает свой водород ПВК. В результате из ПВК образуется молочная кислота, а НАД способен уже принимать новые порции водорода, поддерживая тем гликолиз: СН3-СО-СООН + НАД. H2-^CH3-СНОН-СООН + НАД.
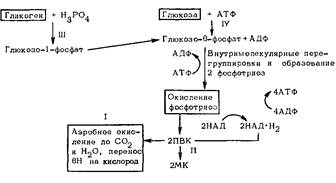
Схема 8. Аэробное и анаэробное окисление глюкозы
MK – молочная кислота. // – аэробное окисление; // – гликолиз; 77/ – путь от гликогена; IV – путь от глюкозы
На первых порах приспособления к гипоксии интенсивность гликолиза возрастает, увеличивается синтез его ферментов. Повышенными возможности гликолиза остаются и при длительном приспособлении к гипоксии. Но у гликолиза три существенных недостатка. Во-первых, аэробно окисляться могут самые различные вещества, а гликолизируется только глюкоза; запасы же гликогена в организме ограниченны. Во-вторых, гликолиз энергетически малоэффективен. В-третьих, организму прежде всего кровь, наводняется молочной кислотой; положение ухудшается еще и тем, что в результате неполного окисления жирных кислот из них образуется в-оксимасляная и ацетоуксусная кислоты, сдвигающие, как и молочная кислота, реакцию внутренней среды в кислую сторону и способные превращаться в ядовитый для организма ацетон.
Естественно, что сдвиг реакции среды в кислую сторону для организма неблагоприятен: если он велик, многие ферменты будут работать не в условиях оптимума кислотности, а значит, проявлять кемаксимальную активность: внутри молекул многих белков могут нарушиться водородные и ионные связи, в результате чего произойдет снижение или даже утрата функциональных свойств данных белков. Конечно, у организма есть средства противостоять этому. Кровь и ткани обладают резервной щелочностью, которая может регулировать реакцию их среды, что осуществляется буферными системами, которые способны связывать при изменении реакции в кислую сторону и OH – при сдвиге в щелочную. Примером может служить бикарбонатнэя буферная система: при избытке H+ бикарбонат, присоединяя их, переходит в плохо диссоциирующую угольную кислоту, а при избытке ОН» угольная кислота превращается в ее двууглекислую соль, а ее водород, соединяясь с гидроксидом, образует крайне слабо диссоциирующую воду. Таким же действием обладают и другие буферные системы: фосфатная, состоящая из одно- и двузамещенной солей фосфорной кислоты, и белки плазмы крови, поскольку они имеют кислую группу – карбоксил и щелочную аминогруппу. Первая связывает ОН-, образуя воду, и переходит в COO-, а вторая – H+, прекращаясь в NH3». Буферным веществом является также и гемоглобин. На долю бикарбонатов приходится 53% всей буферной емкости крови, на долю фосфатов – 5%, белков – 7, гемоглобина 35%. Таким образом, буферные вещества компенсируют сдвиги реакции среды в кислую или щелочную сторону, убирая, связывая ионы H и ОН.
Так как на высотах понижено парциальное давление не только кислорода, но и углекислоты, организм при усиленном дыхании теряет ее в повышенных количествах. Это тоже уменьшает кислотность крови и может привести даже к сдвигу реакции ее в щелочную сторону. Но потери углекислоты вызывают падение возбудимости дыхательного центра в мозгу и ослабление дыхания. Буферные же системы имеют определенную емкость, и она может быть исчерпана. Реакция снова сдвигается в кислую сторону, что называют некомпенсированным ацидозом.
Для борьбы с избытком молочной кислоты организму остается возможность в малой части выделить ее с мочой, а все остальное – химически устранить, что происходит двумя путями. Первый – окисление молочной кислоты до CO2 и Н2О в печени и мышце сердца, но для этого необходимо повышенное поглощение кислорода, что исключено во время пребывания на высоте и возможно лишь при возвращении на более низкие уровни. Второй путь – гликонеогенез, т.е. переработка молочной кислоты в глюкозу и гликоген. Этот цикл реакций, названный по фамилии открывшего его американского биохимика К. Кори циклом Кори, предоставляет орга-ёнтзму ряд преимуществ. Так, устраняется не только молочная кислота, но и некоторое количество H+, присоединяемое к продуктам превращения молочной кислоты в глюкозу, а также увеличивается снабжение центральной нервной системы необходимой ей глюкозой. Таким образом, цикл Кори – один из важных приспособительных механизмов энергетического обмена.
Но длительное пребывание на высотах и тем более постоянное обитание там требует от организма долговременной адаптации к гипоксическим условиям. Сущность и молекулярные механизмы этого приспособления можно понять, сопоставив биохимические и физиологические особенности организма людей, живущих на уровне моря и акклиматизировавшихся к высотам при эпизодическом пребывании там, с постоянно живущими в горах.
Если житель равнины сразу поднимется на значительную высоту, он будет там чувствовать себя весьма неуютно, у него может развиться горная болезнь. Если же высота будет осваиваться постепенно или пребывание на ней будет достаточно длительным, то наступит акклиматизация, человек будет чувствовать себя на высоте достаточно комфортно и сможет выполнять там значительные физические нагрузки. Высотный потолок его возрастет, т.е. снижение насыщения крови кислородом и те изменения функций, которые наступали на данной высоте до акклиматизации, теперь проявятся только на значительно большем уровне. Наконец, у аборигенов гор наибольший высотный потолок, и в тех условиях, к которым жителям равнин надо акклиматизироваться, у них не наблюдается никакого повышения и напряженности физиологических функций, например у непальских шерпов и жителей южноамериканских Анд. Такие же закономерности обнаруживаются при сопоставлении горных животных с обитающими на уровне моря, а также акклиматизированными к высоте.
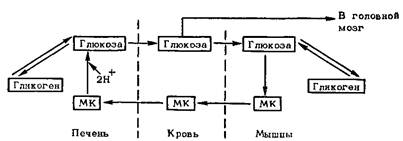
Схема 9. Цикл Кори
В чем же молекулярная и функциональная основа этих отличий? Все приспособления, происходящие на молекулярном уровне, можно разделить на три группы: облегчающие перенесение организмом недостатка кислорода, направленные на борьбу за возможно большее поглощение кислорода в гипоксических условиях и обеспечивающие наиболее эффективное использование кислорода, поступившего в легкие.
К первой группе относятся повышение содержания в мышцах КФ и активности креатинкиназы – одного из основных ферментов, обеспечивающих ресинтез АТФ. При этом на 20–30% увеличивается содержание гликогена в мышцах, а возможности гликолиза – на 40–100%; почти на 30% возрастает резервная щелочность крови. Все это создает лучшие условия для поддержания уровня АТФ в тканях и для сохранения постоянства реакции крови в условиях гипоксии.
При приспособлениях второй группы увеличивается возможность поглощения кислорода в легких прежде всего за счет большего объема крови, проходящей через них в единицу времени. У адаптированного к высоте человека он на 25% превышает величины, характерные для жителей равнин. Кроме того, возрастает кислородная емкость организма за счет повышения объема циркулирующей крови, количества эритроцитов в ней, созревание которых в костном мозгу стимулируется гипоксией, и содержания гемоглобина. При этом не изменяется объем плазмы крови, составляя и у жителей равнин, и у горцев около 45 мл на 1 кг массы тела, а объем эритроцитов у первых равен 35 мл/кг, а у вторых – 65 мл/кг. В горах возрастает и содержание гемоглобина в крови. Так, по данным одной из гималайских экспедиций, 2-недельное пребывание человека на высоте 4000–5000 м приводит к увеличению содержания гемоглобина на 32%, а на высоте 5800 м – на 41%. При акклиматизации крыс в барокамере с атмосферным давлением, соответствующим высоте 9000 м, гемоглобина в крови становится больше на 38%. У животных, постоянно обитающих в горах, оно существенно выше, чем у живущих на уровне моря: у горных сусликов, например, на 55% выше, чем у равнинных. То же можно сказать и о человеке. У жителей Памира содержание гемоглобина на 20% выше, чем у жителей равнины, а у жителей Анд – на 30%. Такие же изменения вызывает приспособление к гипоксии и в отношении миоглобина. Синтез его возрастает, а содержание в скелетных мышцах и миокарде увеличивается, повышая возможности депонирования кислорода в организме. Акклиматизация крыс к высоте 3810 м сопровождается повышением содержания миоглобина в их мышцах на 50 – 70%; у собак, обитающих на высоте 3730–4510 Mf на 40 – 70% более значительно, чем у живущих на равнине. Интересные данные в этом отношении дает сопоставление равнинных и горных животных. Как видим, у вторых оно значительно выше и в мышце сердца, и в скелетной мышце.
Приспособление к недостатку кислорода необходимо не только человеку и животным, живущим в горах, ко и многим видам, ведущим водный образ жизни или хотя бы совершающим периодические ныряния с пребыванием какое-то время под водой. Например, водные млекопитающие могут находиться под водой от 10–15 до 50 мин, а водные черепахи – до нескольких часов. У тех и других содержание миоглобина выше, чем у наземных животных. Так, у тюленей и китов оно достигает 16 – 40 г. на 100 г. высушенной ткани. Если сравнить человека и тюленя с одинаковой массой тела, то запас кислорода, связанного с миоглобином, у первого равен 335 мл, а у второго – 2530 мл. Это почти на порядок больше! В мышцах неныряю щи ч животных миоглобина значительно меньше, чем у ныряющих, но близких им.
Миоглобин обладает намного большим сродством к кислороду, чем гемоглобин крови, и легко присоединяет его, забирая от гемоглобина. Вместе с тем он легко отдает его клеточным окислительным системам, поддерживая окислительные процессы. При приспособлении организма к высотной гипоксии возможности этого возрастают. Миоглобин не только поглощает, резервирует и отдает кислород, но и осуществляет в клетке направленный перенос его к окислительным ферментам, активируя их.
Наконец, третья группа приспособительных изменений, направленных на более эффективное использование кислорода, находит выражение в увеличении количества митохондрий, активности и содержания ряда окислительных ферментов. Активность НАД* H2-цитохром-оксидоредуктазы возрастает на 15%, сукцинатдегидрогс-назы – на 85, цитохромоксидазы – на 100%. Содержание цитохрома с – одного из важных компонентов дыхательной цепи – в разных органах повышается на 16 – 300%. Возрастает и эффективность окислительного фосфорилирования, т.е. выход АТФ на единицу поступившего в митохондрии кислорода; на 30% увеличивается коэффициент Р/О. При этом повышается не только скорость окислительного фосфорилирования, но и активность АТФазы, т.е. возможность и использования АТФ, и возобновления ее запасов, в результате чего того же количества АТФ оказывается достаточно для выполнения большего объема функциональной работы. Увеличение эффективности процессов аэробного окисления наблюдается как у рептилий, так и у млекопитающих животных, не только ведущих водный образ жизни, но и совершающих ныряния. В их мышцах повышено содержание многих компонентов дыхательной цепи, в частности цнто-хрома с.
В результате произошедших молекулярных изменений организм, приспособленный к высотам, в условиях сниженного парциального давления или содержания кислорода во вдыхаемом воздухе потребляет больше кислорода. Так, если в нормальных условиях равнины во вдыхаемом воздухе 21% кислорода, то на высоте его меньше: до 12%; газообмен у не приспособленных к гипоксии животных уменьшается на 11%, в то время как у приспособленных он не изменяется. При еще большем снижении содержания кислорода потребление его у последних выше, чем у первых, на 100–120%. Возрастает и устойчивость высших структур клеточных белков к повреждающему действию гипоксии. Об этом можно судить по способности клеток к прижизненной окраске безвредными красителями: чем больше повреждение структуры белка, тем интенсивнее он окрашивается. Такие исследования показали, что при выдерживании в гипокеических условиях клеточные белки не адаптированных к ним животных на 31% интенсивнее окрашиваются, чем приспособленных.
Но известны животные, которые могут одинаково нормально жить и при хорошей доступности кислорода, и при крайне низком содержании его, и почти при полном отсутствии, и даже такие, которым кислород не только не нужен, но даже вреден. Первых называют анаэробами факультативными, вторых – облигатными. К первым относятся водные черепахи и многие рыбы, ведущие придонный образ жизни. Дело в том, что в придонной воде содержание кислорода может доходить до 15% той величины, которая наблюдается при насыщении воды воздухом.
В ряде случаев в придонных слоях может и не быть кислорода. Вместе с тем установлено, что в придонной бескислородной зоне оз. Танганьика обитают 10 видов рыб. Факультативными анаэробами являются, например, карп и серебряный карась. В зимнее время, когда реки и пруды покрыты льдом, они на 2 3 мес лишены кислорода.
Для факультативных анаэробов характерны очень высокие возможности гликолиза, устойчивость к накоплению в организме больших концентраций молочной кислоты и наличие дополнительных путей генерирования АТФ бескислородным механизмом. Наблюдения, проведенные на водных черепахах, показали, что в первые 30 мин после погружения в воду интенсивность аэробного окисления у них не изменяется, но напряжение кислорода в легких и крови быстро падает. В следующие 30 мин интенсивность обмена снижается до 40% от исходного, а кислородные запасы иссякают. Затем в течение нескольких часов черепаха живет исключительно за счет энергии гликолиза, а содержание молочной кислоты в крови и тканях достигает 60 ммоль и более. Образовавшийся же избыток молочной кислоты устраняется с помощью окисления лишь при возвращении черепахи в воздушную среду. Заметим, что в крови человека и наземных животных молочной кислоты не больше 18–20 ммоль/л, и то при мышечной деятельности максимальной или субмаксимальной интенсивности.
Кроме гликолиза у факультативных анаэробов существуют другие пути анаэробного генерирования АТФ, связанные с декарбоксилированием б-кетоглютаровой и пировиноградной кислот, отщеплением их карбоксильных групп и образованием CO2. Эта сложная, многоступенчатая цепь реакций изучена еще недостаточно. Но из всего сказанного следует, что набор ферментов в тканях факультативных анаэробов должен если не качественно, то уж во всяком случае количественно и по характеру регуляции активности существенно отличаться от того, что имеет место у облигатных аэробов, и позволять им черпать энергию и из аэробных, и из анаэробных окислительных процессов.
Облигатные же анаэробы, например кишечные глисты, получают необходимую им энергию исключительно за счет анаэробных реакций. Кислород действует на них убийственно. Не случайно одним из способов изгнания глистов из кишечника является вдувание туда кислорода. Установлено, что у облигатных анаэробов утрачена возможность синтеза ряда ферментов, участвующих в реакциях аэробного окисления.
Но приспособление к недостатку кислорода может идти и по функциональному пути, на органном уровне. В этом отношении интересным примером могут служить антарктические рыбы белокровки. У этих рыб, видимо в результате давно произошедшей мутации, в крови нет ни эритроцитов, ни гемоглобина, а содержание железа в организме в 25 раз ниже, чем у других видов рыб, обитающих в тех же условиях. Бесцветная плазма крови у них переносит к органам только растворенный в ней кислород, концентрация которого очень невелика. Так как при фильтрации воды через жабры в крови рыб не достигается насыщающая ее концентрация кислорода, в процессе эволюции выработался дополнительный механизм – диффузия кислорода через кожу, снабженную мощной сетью капилляров. Кроме того, возможности поглощения кислорода возрастают в связи с высокой интенсивностью кровообращения. Так, сердце белокровок в 3 раза больше, чем у равновеликих им нототений и других антарктических рыб. Но, конечно, у этих рыб более высоки и возможности анаэробного генерирования АТФ.
Приспособление к высоким давлениям
По-иному влияют на организм высокие давления, и иначе происходит приспособление организма к ним. В условиях высокого гидростатического давления главным повреждающим фактором является само давление. Правда, в глубоких водах снижено содержание кислорода, и к этому глубоководные животные должны быть приспособлены. Но по сравнению с эффектом колоссальных давлений влияние этого фактора сравнительно невелико. Конечно, внешнему давлению, которое испытывают глубоководные животные, противостоит такое же по величине внутреннее давление, что препятствует их деформации, расплющиванию. Однако сам факт большого давления остается, а высокие давления повреждают четвертичные и третичные структуры клеточных белков, образованные водородными и ионными связями, и таким образом влияют на скорость химических реакций.
Многие структурные и ферментные белки содержат две или более субъединиц, соединенных между собой иековалентными связями. Например, лактат-дегидрогеназа, отнимающая от молочной кислоты водород, и киназа фосфорилазы, превращающая малоактивную фосфорилазу b в высокоактивную фосфорилазу а, состоят каждая из четырёх субъединиц. Расщепление фермента на субъединицы лишает его активности или резко снижает ее. Из субъединиц построены и сократительные белки мышц – миозин и актин: первый – из пяти субъединиц, а второй – из очень большого числа глобулярных молекул G-актина. Расщепление этих сложных структур высоким давлением и препятствие сборке субъединиц G-актина в фибриллярный F-актин делает невозможным взаимодействие между этими белками и мышечное сокращение. Установлено также, что давление 101 100 кПа, соответствующее глубине 10 км, лишает актомиозин мышц корюшки его ферментативной активности, способности трансформировать химическую энергию АТФ в механическую энергию мышечного сокращения, а у карпа снижает ее на 50%, затрудняет высокое давление и дерепрессию генов, полимеризацию РНК, сборку рибосом в полирибосомы, а следовательно, и синтез белков. Наконец, высокое давление изменяет структуры клеточных мембран и уменьшает их проницаемость.
Влияние высокого давления на скорость биохимических реакций зависит от характера изменения объемов реагирующих веществ в ходе реакции. Если у фермент-субстратного комплекса или конечного продукта больший объем, чем у исходных веществ, то давление ингибирует такую реакцию, а если объем в ходе реакции уменьшается, то активирует ее. Изменения объемов зависят не только от структуры и величины молекул и комплексов, образующихся в процессе реакции, но и от взаимодействия их с молекулами внутриклеточной воды, которые могут вступать в физико-химическую связь и с ферментом, и с фермент-субстратным комплексом, и с продуктом реакции, гидратируя их. Степень гидратации, а значит, и изменение объема зависят от ряда условий: электрического заряда молекул, количества неорганических ионов, рН среды и т.д. Естественно, что все эти обстоятельства не могут не влиять положительно или отрицательно на скорость и направление реакций обмена веществ.
Мы не будем касаться морфологической стороны приспособления особенностей форм и строения глубоководных организмов, так как молекулярная основа этого неизвестна. Да и для выяснения молекулярных основ адаптации организмов к высоким давлениям сделано еще не так много. Прежде всего установлено различие между свойствами белков глубоководных организмов и животных, совершающих значительные вертикальные миграции под водой: у первых белки сохраняют свои высшие структуры, а ферменты оптимально работают только при больших давлениях, к которым они нечувствительны, но претерпевают существенные изменения при понижении давления; у вторых белки не реагируют на большие перепады давления. Чем обусловлены эти свойства, мы пока не знаем. Возможно, что они зависят от особенностей высших структур белков и от каких-то условий внутриклеточной среды, способствующих сохранению этих структур и благоприятному изменению объемов компонентов различных биохимических реакций. Тут может играть роль присоединение к белку каких-то веществ, снимающих ингибирующее влияние высоких давлений. Так, фруктозодифосфат снимает ингибирующее влияние давления на чувствительный к нему фермент – пируваткиназу.
Вторая возможность приспособления обмена веществ к высоким давлениям – выбор альтернативных путей биохимических превращений. Нередко в организме за один и тот же субстрат конкурируют несколько ферментов, направляющих его по разным путям в зависимости от концентрации субстрата, условий среды и т.д. Примером может служить судьба промежуточного продукта гликолиза – глюкозо-6-фосфата, который образуется в результате фосфорилирования глюкозы. Далее превращения его могут идти по одному из четырех путей: он может подвергаться гликолизу до молочной кислоты; окисляться в фосфоглюконат и участвовать в образовании пентоз; использоваться для синтеза гликогена; расщепляться на глюкозу и фосфорную кислоту. Выбор пути зависит от того, какая из четырех реакций наименее связана с давлением. Однако этот вопрос требует дальнейших исследований.
Большую опасность для обитателей глубин представляет перемещение по вертикали. Почти все глубоководные животные при поднятии их на поверхность в тралах оказываются мертвыми. У рыб при этом пищевод выпячивается изо рта, а кишечник – из анального отверстия, глаза выходят из орбит, чешуя отстает и отпадает. Но есть ряд животных, хорошо переносящих такие перемещения, например поднимающиеся на поверхность и быстро ныряющие в глубину акулы, макрели и живущие придонно ракообразные. Известный исследователь морских глубин В. Биб привязывал омара снаружи своей батисферы, подвергая его при погружении разности давлений 10 110 кПа, и омар возвращался живым и неповрежденным. Вообще омары, лангусты и креветки, извлекаемые на поверхность моря, всегда остаются живыми и активными. Что лежит в основе такой адаптации, мы еще не знаем. У рыб же, резко меняющих глубину пребывания, одной из приспособительных особенностей является отсутствие плавательного пузыря, заполненного воздухом, объем которого при быстром поднятии резко расширяется.
Приспособление к температуре среды
Для защиты от температурного фактора в строении многих животных имеются специальные приспособления. Так, у ряда насекомых хорошую термоизоляцию обеспечивает густой покров волосков на грудном отделе: между волосками находится слой неподвижного воздуха, уменьшающий теплоотдачу. Тунцы могут поддерживать температуру своих мышц на 8 – 10 0C выше температуры воды благодаря наличию особых теплообменников – тесного переплетения артериальных и венозных капилляров, на которые распадаются идущие от жабр артерии и от мышц к жабрам вены. Первые несут кровь, охлажденную водой, вторые – согретую работающими мышцами. В теплообменнике венозная кровь отдает тепло артериальной, что способствует сохранению более высокой температуры в мышцах. У водных млекопитающих термоизоляцией служит толстый слой подкожного жира, а у белого медведя, кроме того, и непромокаемая до кожи шерсть. У водоплавающих птиц ту же роль играют перья, покрытые жироподобной смазкой.
О том, как велико значение этой смазки, рассказывает в своих воспоминаниях крупный немецкий зоолог и основатель всемирно известного Гамбургского зоологического сада К. Гакенбек. Он с детства увлекался животными. Однажды отец подарил ему несколько диких уток с подрезанными крыльями, поэтому они улететь не могли. И маленький Карл пустил их плавать в металлический бак. Но бак оказался из-под мазута, в котором утки вымазались с ног до головы. Увидев такой непорядок, мальчик тщательнейшим образом вымыл уток теплой водой с мылом и пустил их плавать в другой, чистый бак. На следующее утро все утки лежали мертвыми на дне: теплая вода и мыло удалили не только мазут, но и всю жировую смазку, в результате чего утки переохладились и погибли.
Мы уже знаем, что гомойотермные животные могут поддерживать температуру тела в гораздо большем диапазоне температур, чем пойкилотермные, однако те и другие гибнут при примерно одинаковых чрезмерно высоких или чрезмерно низких температурах. Но пока этого не произошло, пока температура не достигла критических значений, организм борется за поддержание ее па нормальном или хотя бы на близком к нормальному уровне. Естественно, что в полной мере это свойственно гомойотермным организмам, обладающим терморегуляцией, способным в зависимости от условий усиливать или ослаблять как теплопродукцию, так и теплоотдачу. Теплоотдача – процесс чисто физиологический, он происходит на органном и организменном уровнях, а в основе теплопродукции лежат и физиологические, и химические, и молекулярные механизмы. Прежде всего это озноб, холодовая дрожь, т.е. мелкие сокращения скелетных мышц с низким коэффициентом полезного действия и повышенным образованием тепла. Этот механизм организм включает автоматически, рефлекторно. Эффект его может быть повышен активной произвольной мышечной деятельностью, также усиливающей теплообразование. Не случайно, чтобы согреться, мы прибегаем к движению.
У гомойотермных существует возможность образования тепла и без сокращения мышц. Происходит это в основном в мышцах, а также в печени и других органах следующим образом. При транспорте электронов и протонов по дыхательной цепи энергия окисляемых веществ не рассеивается в виде тепла, а улавливается в форме образующихся макроэрги-ческих соединений, обеспечивающих ресинтез АТФ. Эффективность этого процесса, открытого выдающимся биохимиком В.А. Энгельгардтом и получившего название дыхательного фосфорилирования, измеряется коэффициентом Р/О, показывающим, сколько атомов фосфора было включено в АТФ на каждый атом использованного митохондриями кислорода. В обычных условиях в зависимости от того, какое вещество окисляется, этот коэффициент разен двум или трем. При охлаждении организма окисление и фосфорилирование частично разобщаются. Та пли иная часть окисляемых веществ вступает на путь «свободного» окисления, в результате чего уменьшается образование АТФ и повышается выделение тепла. При этом, естественно, коэффициент Р/О понижается. Разобщение это достигается действием гормона щитовидной железы и свободными жирными кислотами, в повышенных количествах поступающими в кровь и приносимыми ею к мышцам и другим органам. При увеличении внешней температуры, наоборот, сопряжение окисления и фосфорилирования усиливается, а теплопродукция снижается.
Кроме мышц и печени, для которых теплообразование служит не основной, а побочной функцией, в организме млекопитающих животных есть и специальный орган теплопродукции – бурая жировая ткань. Она располагается около сердца и по пути крови к жизненно важным органам: сердцу, мозгу, почкам. Клетки ее исключительно богаты митохондриями, и в них очень интенсивно идет окисление жирных кислот. Но оно не сопряжено с фосфорилированием АДФ, а энергия окисляемых веществ выделяется у них 'в виде тепла. Усилителем окислительных процессов в бурой жировой ткани является адреналин, а разобщителем дыхания и фосфорилирования – образующиеся в ней в больших количествах жирные кислоты.
Интересный механизм поддержания температуры мышц был недавно открыт у шмелей известным английским биохимиком Э. Ньюсхолмом. У всех животных образующийся в процессе гликолиза фруктозофосфат, присоединяя от АТФ еще одну частицу фосфорной кислоты, превращается во фруктозодифосфат, который направляется далее на путь анаэробного окисления. У шмелей же он расщепляется на фруктозо-6-фосфат и фосфорную кислоту с выделением тепла: Ф-6-Ф + АТФ -> ФДФ + АДФ; ФДФ -> Ф-6-Ф + К3РО4 – f – тепло, что в сумме дает реакцию АТФ –>-АДФ -\~ Н3РО4 + тепло. Дело в том, что в противоположность другим животным у шмелей фруктозодифосфа-таза не угнетается продуктами расщепления АТФ. В результате шмели достигают разности температур между мышцами и окружающей средой порядка 8–20 °С, что позволяет им активно передвигаться и кормиться в прохладную погоду, неблагоприятную для других насекомых.
В экстренном приспособлении к изменениям температурного режима у гомойотермных важную роль играют и гормоны. В условиях низких температур в кровь выбрасывается повышенное количество адреналина, стимулирующего мобилизацию глюкозы и жирных кислот и интенсивность окислительных процессов. В крови происходит освобождение глюкокортикоидов от связи с белками, а затем и новое поступление их в кровь из коры надпочечников. Они повышают чувствительность периферических адренорецепторов, усиливая тем действие адреналина. Активируется деятельность щитовидной железы, гормоны которой вызывают частичное разобщение дыхания и фосфорилирования в митохондриях мышц и печени, увеличивая теплообразование. При действии высоких температур интенсивность окислительных процессов и теплообразование снижаются, возрастает теплоотдача. Но все это хорошо для экстренного, кратковременного приспособления организма и было бы даже вредным для него при длительном изменении температурных условий. Действительно, если бы животные, обитающие в области низких температур, защищались от них, например, только холо-довой дрожью, неизвестно, как они могли бы вести активную жизнь, добывать пищу, спасаться от врагов и т.п. Значит, при длительной адаптации к той или иной температуре приспособительные механизмы должны быть иными: обеспечивать нормальное существование организма в этих условиях.
Для того чтобы произошла химическая реакция, должны возникнуть напряжение или деформация и ослабление связей в молекулах реагирующих веществ. Необходимую для этого энергию называют энергией активации. Повышение температуры на 10 0C увеличивает скорость реакции в 2–3 раза за счет возрастания числа активированных молекул. При понижении температуры наблюдаются изменения обратного порядка. Если бы организм строго следовал этому закону, то при изменении температуры среды он оказался бы в весьма трудном положении: низкие температуры настолько замедлили бы реакции обмена веществ, что жизненные функции не могли бы протекать нормально, а при высоких температурах они чрезмерно ускорились бы. На деле же мы видим совсем другое. Так, у рыб, приспособленных к высоким и низким температурам, различия в интенсивности обмена веществ не очень велики и вполне соизмеримы. Иначе говоря, реакции обмена веществ у этих видов имеют разный температурный оптимум. Например, у енотовидной собаки интенсивность обмена веществ наиболее низка при 15 °С, а в обе стороны от этой точки она возрастает. Температура же тела в амплитуде 35 0C сохраняется почти постоянной. А это значит, что температурные условия протекания реакций обмена веществ в этом диапазоне сохраняются оптимальными. При сопоставлении двух близких видов животных, но обитающих в разных условиях, мы видим, что у песца постоянство интенсивности обмена веществ и температуры тела в большом диапазоне температур среды намного лучше выражено, чем у лисы. Что интересно: интенсивность обмена веществ при понижении окружающей температуры не уменьшается, а сохраняется на постоянном уровне или повышается, тогда как, согласно химическим законам, должно было бы быть наоборот. Такая возможность открылась перед живыми организмами потому, что все реакции обмена веществ ферментативны. А суть действия ферментов в том, что они резко снижают энергию активации реагирующих молекул. Кроме того, в зависимости от условий среды они могут изменять ряд своих свойств: каталитическую активность, оптимум температуры и кислотности, степень сродства к субстрату. Поэтому причины способности организма «уклоняться» от химических законов следует искать в изменениях ферментных белков.
Изменения эти в связи с приспособлением к температурному фактору могут идти по трем путям: увеличивать или уменьшать число молекул данного фермента в клетке, изменять набор ферментов в ней, а также свойства и активность ферментов. Первый путь имеет свои резоны. Ведь любая молекула фермента в каждый данный момент может взаимодействовать с одной молекулой субстрата. Поэтому, чем больше молекул фермента в клетке, тем значительнее будет выход продуктов реакции, а чем меньшим оно будет, тем ниже и выход. Это в какой-то мере может компенсировать температурное снижение или повышение интенсивности обмена веществ. Но эта компенсация ограничивается и возможностью синтеза ферментов, и пространственными соображениями. Клетка те может вместить слишком большое количество новых макромолекул ферментов. Тем не менее имеются уже твердо установленные данные, что при приспособлении к холоду возрастают активность и содержание в мышцах таких важнейших ферментов аэробного окисления, как сукцинатдегидрогеназа и цитохром-оксидаза.
Несомненно, что более эффективен второй путь, определяемый репрессией синтеза одних ферментов и индукцией синтеза других, В условиях низких температур повышенно синтезируются ферменты, в большей степени снижающие энергию активации, а в условиях высокой – менее значительно влияющие на нее. Это относится прежде всего к изоферментам. У фермента лактатдегидро-геназы пять изоформ. При этом изофермент II4 более значительно снижает энергию активации, чем M4. Поэтому при приспособлении к низким температурам в большей степени синтезируется первый, а к высоким – второй. У фермента нервной системы холинэстеразы две изоформы с разными возможностями снижения энергии активации. Исследование мозга радужной о^орели, приспособившейся к разным температурным условиям, показало, что при адаптации к температуре 2 0C присутствует исключительно изофермент I, к температуре 17 0G – изофермент II, а у живущих при 12 0G – обе изоформы. Это касается и сезонных изменений: зимой синтезируется преимущественно изоформа I, а летом – изоформа II.
Третий путь приспособления – прежде всего изменение сродства фермента к субстрату. В основе этого – изменения высших структур ферментных белков и свойств их активных центров. При этом способность их связывать субстрат, образуя фермент-субстратный комплекс, повышается или понижается. Непосредственными причинами перестроек являются изменение электростатических свойств активного центра, степень диссоциации атомных группировок, принимающих участие в связывании субстрата, ионное окружение активного центра и изменение его пространственной формы. Сдвиги температурной зависимости активности ферментов могут быть обусловлены и присоединением к их молекулам различных аллостерических эффекторов: белков, фосфолипидов, неорганических ионов и др. У серебряных карасей, приспособленных к обитанию при 5 и 25 °С, исследовали активность сукцииатдегидрогеназы, встроенной в мембраны митохондрий и связанной с фосфолипидами. Фермент выделили в чистом виде и освободили от фосфо-липидов. У обеих групп строение его было совершенно одинаковым, а активность – ниже максимальной. Затем к ферментному белку добавляли фосфолипиды из «Холодовых» и «тепловых» митохондрий. Первые активировали фермент больше, чем вторые. Анализ фосфолипидов показал, что в Холодовых митохондриях жирные кислоты фосфолипидов наиболее насыщенные. Возможно, что в этом причина снижения степени сопряжения дыхания и фосфор при приспособлении к холоду и повышение его при адаптации к высоким температурам.
Приспособление к температурным условиям не ограничивается только изменениями в области ферментных систем, хотя они и являются основой. При адаптации к низким температурам в мышцах возрастает содержание КФ, а в жировых депо – резервного жира, служащего как высокоэффективным источником энергии, так и термоизолятором. В фосфолипидах клеточных мембран увеличивается содержание ненасыщенных и полиненасыщенных жирных кислот, что препятствует их затвердеванию при низких температурах. Наконец, у животных, способных к перенесению очень низких температур, в крови, тканевых жидкостях и клетках обнаружены биологические антифризы, препятствующие замерзанию внутриклеточной воды. Впервые они были выделены у антарктических рыб – нототении и трематомуса. По своей природе они гликопротеины, т.е. соединения сахара галактозы с белком. Связывающим звеном является азотосодержащее основание ацетп л га лактозам ии. Их MM может достигать 21 500, и для них характерно высокое содержание гид роке ильных групп, уменьшающих возможности взаимодействия между молекулами воды и образование льда. Чем с более низкими температурами встречается организм, тем выше содержание антифриза. Летом оно меньше, зимой больше. У арктических насекомых роль антифриза выполняет глицерин, тоже богатый гидроксильными группами. В гемолимфе и тканях этих животных содержание глицерина возрастает с понижением температуры.
Закаливание
Закаливание – приспособление человека к низким температурам, с которыми мы встречаемся повседневно как в быту, так и в труде. Закаливание весьма желательно и даже необходимо человеку по двум причинам. Во-первых, наши отдаленные предки обитали в условиях жаркого климата, а поэтому человек – теплолюбивый организм. Во-вторых, на протяжении всей истории человечества оно искусственно создавало себе оптимальный микроклимат, используя одежду, жилище, отопление, в результате чего границы эффективной теплорегуляции в организме человека сузились по сравнению с тем, что свойственно животным. Закаливание эти границы расширяет.
Первые эксперименты по закаливанию были проведены А. Назаровым в лаборатории В.В. Пашутина более ста лет тому назад. А. Назаров установил, j что если собаку погружать в ванну с температурой воды 10 0C, то в первые дни температура тела у нее снижается на 6 – 7 °С, а после семи–восьми погружений она становится стабильной, колеблясь в пределах ±0.3 °. Дальнейшие исследования многочисленных авторов показали, что под влиянием действия холода значительно возрастают потребление кислорода и интенсивность окислительных процессов. Например, при купании в Неве осенью, зимой и весной – в 2 – 3 раза, а по мере приспособления организма – всего на 20–30%. Но зато повышение интенсивности и эффективности окислительных процессов в промежутках между охлаждениями остается постоянным, что создает лучшую энергетическую базу для все время идущего самообновления и синтеза белков. Необходимо только, чтобы охлаждение организма с первых шагов не было слишком сильным, оно должно нарастать постепенно.
Очень хороший эффект дает сочетание охлаждений с мышечной деятельностью. На значение мышечной деятельности для приспособления организма к холоду указывал еще в прошлом веке видный русский физиолог И.Р. Тарханов. А вот что дали исследования, проведенные уже в наше время. Две группы испытуемых занимались утренней гимнастикой круглый год на открытом воздухе: одна – всегда обнаженной до пояса, а вторая – сначала в тельняшках, а при сильных морозах – в пальто. Гимнастическая нагрузка у обеих групп была одинаковой. В первой группе испытуемых даже при морозе –30 0C занятия в полуобнаженном виде не вызывали неприятных ощущений холода и снижения температуры тела, а во второй по сути дела никакого закаливания не наступило, и температура тела всякий раз снижалась. Интерес этих экспериментов не только в том, что сочетание воздействия холода с физической нагрузкой приводило к высокому закаливающему эффекту, но и в том, что сама по себе физическая нагрузка такого эффекта не оказывала.
Хороший результат дает и сочетание Холодовых процедур с тепловыми: несколько заходов в сауну с температурой более 100 °С, чередуемое всякий раз с окунанием в прорубь или обтиранием снегом с ног до головы. Резкие переходы от ограничения теплопродукции и увеличения теплоотдачи к стимулированию первой и уменьшению второй эффективно тренируют и систему терморегуляции, и динамику кровеносных сосудов кожи. К тому же это просто приятно.
Закаливание разумно начинать с детского возраста: хождение летом босиком; сначала прохладные, потом все более холодные обтирания или обливания; летом купание при температуре воды от 17 0C и выше; затем постепенный переход по мере взросления к методам закаливания, применяемым взрослыми. Не следует в холодную погоду излишне кутать ребенка, одежда должна быть разумно, но не чрезмерно теплой и не стесняющей движений. И так – до старости, так как стимуляция окислительных процессов Холодовыми процедурами эффективно поддерживает синтез белков тканей, замедляя тем процесс старения, в основе которого лежит прежде всего снижение синтеза белков.
© 2009 База Рефератов