
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Реферат: Роль эндофрина в нейроэндокринной регуляции функций иммунной системы
Реферат: Роль эндофрина в нейроэндокринной регуляции функций иммунной системы
На правах рукописи
Гейн Сергей Владимирович
РОЛЬ b-ЭНДОРФИНА В НЕЙРОЭНДОКРИННОЙ РЕГУЛЯЦИИ ФУНКЦИЙ ИММУННОЙ СИСТЕМЫ
14.00.36 Аллергология и иммунология
Автореферат диссертации на соискание ученой степени доктора медицинских наук
Пермь - 2007
Работа выполнена в аналитической лаборатории Института экологии и генетики микроорганизмов УрО РАН, Пермь.
Научный консультант:
академик РАН и РАМН, д.м.н., профессор
Черешнев Валерий Александрович
Официальные оппоненты:
доктор медицинских наук, профессор Сибиряк Сергей Владимирович,
доктор медицинских наук, профессор Кузнецов Валериан Фёдорович,
доктор медицинских наук, профессор Юшков Владимир Викторович
Ведущая организация: Институт экспериментальной медицины РАМН, Санкт-Петербург.
Защита состоится «__»__________ 2007 г. в ____ часов на заседании диссертационного совета Д 004.019.01 в Институте экологии и генетики микроорганизмов УрО РАН по адресу: 614081, г. Пермь, ул. Голева, 13. Факс (342)2446711.
С диссертацией можно ознакомиться в библиотеке Института экологии и генетики микроорганизмов УрО РАН, Пермь.
Автореферат разослан «___» _______________2007 г.
Ученый секретарь диссертационного совета,
чл.-корр.РАН Ившина Ирина Борисовна
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Поддержание внутреннего гомеостаза определяется взаимодействием нервной, эндокринной и иммунной систем организма. Известно, что иммунная система многокомпонентна, ее функционирование обеспечивается сложной сетью взаимосвязанных сигналов. Одними из важнейших посредников во взаимодействии нервной и иммунной систем выступают эндогенные опиоидные пептиды, представляющие собой группу факторов и играющих ключевую роль в процессах адаптации организма (Корнева, Шхинек, 1988; Зозуля, Пшеничкин, 1990; Корнева, 2007; Maier, 2003; Pruett, 2003; Bodnar, Klein, 2006; Sharp, 2006; Wilbert-Lampen et al., 2007). Несмотря на то, что в последние годы изучению влияния эндогенных опиоидных пептидов на процессы регуляции иммунитета в литературе уделяется достаточно много внимания (Panerai, Sacerdote, 1997; Tomassini et al., 2003, 2004; Sacerdote, 2003), вопрос о механизмах реализации эффектов биорегуляторных пептидов данного класса остается крайне актуальным.
Основной источник опиоидных пептидов в организме - центральная нервная система. Основная группа пептидных гормонов (адренокортикотропный гормон, b-липотропин, меланоцитстимулирующий гормон, b-эндорфин) образуется в результате расщепления большой молекулы – предшественника проопиомеланокортина. При этом наиболее активным и полифункциональным представителем пептидов группы проопиомеланокортина является b-эндорфин. Основной источник b-эндорфина в центральной нервной системе – аркуатное ядро гипоталамуса, на периферии – промежуточная доля гипофиза, из которой пептид секретируется в кровь при стрессе, шоке, травмах и физических нагрузках. b-эндорфин является ключевым фактором, осуществляющим контроль стрессиндуцированных изменений иммунитета со стороны эндогенной опиоидной системы (Зозуля, Пшеничкин, 1990; Pedersen, Hoffman-Goetz, 2000). Роль других соединений из семейства эндогенных опиоидных пептидов, в частности энкефалинов, при стрессе значительно более скромна, а по мнению отдельных авторов (Owens, 1987) вообще отрицается.
Известно (Panerai, Sacerdote, 1997), что изменение концентрации b-эндорфина в головном, спинном мозге и гипофизе часто сочетается с неврологическими и аутоиммунными нарушениями (мигрень, рассеянный склероз, болезнь Крона). Важную роль b-эндорфин играет в патогенезе инфекционных заболеваний, модулируя функции клеток адаптивного и естественного звеньев иммунной системы при их контакте с микроорганизмами и вирусами (Ляшев, 2000; Plotnikoff, 1999; Sitte et al., 2007). В связи с этим изучение роли b-эндорфина в регуляции иммуногенеза представляет большой интерес.
Широкий спектр биологической активности b-эндорфина определяется его способностью взаимодействовать с различными по своей природе сайтами связывания, к которым относятся опиоидные (налоксон-чувствительные) и неопиоидные (налоксон-нечувствительные) рецепторы. Экспрессия опиатных рецепторов трёх основных классов (µ, d, k) и неопиоидного рецептора на клетках различных органов и тканей, в том числе и клетках иммунной системы, доказана методами радиолигандного связывания и детекции соответствующей РНК (Наволоцкая и др., 2004; Madden, 1995; Plotnikoff, 1999; Bidlack, 2000; Kraus et al., 2006; Lotsch et al., 2006; Sharp, 2006). Подобное распределение участков связывания b-эндорфина объясняет широкий спектр активности данного пептида и указывает на возможность как прямого, так и опосредованного влияния на формирование иммунного ответа (Зозуля, Пшеничкин 1990; Bidlack, 2000; Stanojević et al., 2006).
Нерешенными остаются вопросы, касающиеся изучения роли эндогенной опиоидной системы в стрессиндуцированных изменениях гуморального и клеточноопосредованного иммунитета; различных рецепторов в регуляции выраженности иммунных процессов, индуцируемых эндогенными опиоидами при стрессе, а также на фоне введения двух основных стрессреализующих факторов – глюкокортикоидов и катехоламинов. Малоизученными остаются молекулярные и клеточные механизмы иммунорегуляторного действия b-эндорфина, связанные с эффектами данного гормона на процессы пролиферации, кооперации и дифференцировки клеток иммунной системы, продукцию ряда ключевых цитокинов (g-IFN, IL-4, IL-10, IL-12), являющихся маркерными для регуляторных Т-лимфоцитов 1 и 2 типа (Th1/Th2) и определяющих выбор типа иммунного ответа. В литературе имеются довольно противоречивые данные о влиянии b-эндорфина на функции клеток естественного иммунного ответа (Van den Bergh et al., 1994; Peterson et al., 1998; Voccarino, Kastin, 2000; Sacerdote, 2003; Bodnar, Klein, 2006).
Цель настоящей работы – изучение роли опиатергических механизмов в нейроэндокринной регуляции иммуногенеза с оценкой эффектов b-эндорфина на процессы пролиферации, дифференцировки и кооперации клеток иммунной системы.
Основные задачи исследования
1. Изучить роль основных компонентов эндогенной опиоидной системы в регуляции процессов иммуногенеза в условиях острого стресса.
2. Исследовать возможность опосредованности иммунорегуляторных эффектов глюкокортикоидов и катехоламинов через взаимодействие с эндогенной опиоидной системой.
3. Оценить влияние b-эндорфина на функции клеток адаптивного иммунитета и исследовать механизм действия пептида на процессы пролиферации, кооперации и Th1/Th2-дифференцировки лимфоцитов.
4. Изучить роль b-эндорфина в регуляции функций клеток естественного иммунитета.
Научная новизна работы. Экспериментально обоснована интегральная роль эндогенной опиоидной системы в нейроэндокринной регуляции иммуногенеза в норме, в условиях стрессорного воздействии и при введении стрессреализующих гормонов – глюкокортикоидов и катехоламинов. Впервые изучено влияние b-эндорфина на процессы активации, пролиферации, кооперации и Th1/Th2-дифференцировки Т-лимфоцитов с оценкой роли различных клеточных фракций. Выявлен характер участия опиатных рецепторов различных типов в регуляции иммунных реакций под воздействием b-эндорфина и синтетических лигандов опиатных рецепоров пептидной природы. В экспериментах in vivo и in vitro подтверждено, что основной мишенью b-эндорфина является гуморальное звено иммунного ответа, при этом выявлена зависимость эффектов b-эндорфина от фазы развития иммунной реакции. В модели пролиферативного ответа лимфоцитов обнаружена зависимость эффекта b-эндорфина и селективных агонистов m- и d-рецепторов от концентрации митогена и концентрации исследуемых пептидов. Выявлена ключевая роль d-рецепторов в реализации стимулирующего эффекта b-эндорфина на пролиферацию и продукцию IL-4. Впервые установлена зависимость стимулирующего эффекта опиоидных пептидов на пролиферацию лимфоцитов и продукцию IL-4 от присутствия моноцитов в клеточной культуре. Обнаружено, что b-эндорфин снижает степень выраженности реакции бласттрансформации лимфоцитов при удалении моноцитов из клеточной культуры. Показано, что b-эндорфин стимулирует фагоцитарную активность эффекторов естественного иммунитета и оказывает модулирующее влияние на цитокинпродуцирующую функцию моноцитов и нейтрофилов. Изучен характер участия опиатных рецепторов в регуляции этих процессов.
Теоретическая и практическая значимость. Полученные данные расширяют представление о роли b-эндорфина и опиатных рецепторов разных типов в регуляции иммуногенеза. Обосновано, что формирование иммунного ответа в норме и при стрессе, а также регуляция адаптивного и естественного иммунного ответа реализуется за счёт механизмов, связанных с синтезом опиоидных пептидов. Выявлен механизм влияния опиоидных пептидов на процессы Th1/Th2-дифференцировки Т-лимфоцитов. Полученные экспериментальные данные подтверждают целесообразность использования иммуномодулирующих свойств b-эндорфина в терапии ряда аутоиммунных заболеваний (рассеянный склероз, болезнь Крона и пр.). В связи с широким использованием агонистов опиатных рецепторов в практической медицине, необходим учет последствий их применения. Результаты работы используются в лекционном курсе «Экспериментальная иммунопатология и иммунотерапия» на кафедре микробиологии и иммунологии Пермского государственного университета (614990, Пермь, Букирева 15).
Основные положения, выносимые на защиту
1. В условиях блокады опиатных рецепторов выявляется выраженная модификация иммунорегуляторных эффектов ротационного стресса, глюкокортикоидов и катехоламинов.
2. b-эндорфин в высоких дозах угнетает, в низких - стимулирует гуморальное звено иммунитета, и при этом не влияет на выраженность реакции гиперчувствительности замедленного типа. В клеточных культурах b-эндорфин стимулирует пролиферацию лимфоцитов и продукцию IL-4, не влияет на синтез IL-2 и IFN-g, при этом усиливает степень поляризации Т-хелперов в направлении Th2-клеток. В реализации стимулирующих эффектов b-эндорфина на пролиферацию и продукцию IL-4 доминирующая роль принадлежит d-рецепторам.
3. Клетки моноцитарно-макрофагального ряда играют важную регуляторную роль в направленности эффектов b-эндорфина в отношении функциональной активности CD4+ лимфоцитов.
4. b-эндорфин стимулирует фагоцитарную активность эффекторов естественного иммунитета и оказывает модулирующее влияние на цитокинпродуцирующую функцию моноцитов и нейтрофилов.
Связь работы с крупными программами. Работа проводилась в течение 2000-2007 гг. в соответствии с планом НИР ИЭГМ УрО РАН (номер госрегистрации темы НИР 01.9.009927); в рамках Программы фундаментальных исследований Президиума РАН «Молекулярная и клеточная биология»; гранта РФФИ 06-04-49001, а также грантов молодых учёных Президиума УрО РАН 2003, 2005 гг.
Апробация работы. Материалы диссертации доложены и обсуждены на Международном симпозиуме «Взаимодействие нервной и иммунной систем в норме и патологии», Санкт-Петербург, 2007; V-VIII конференциях с международным участием «Дни иммунологии в Санкт-Петербурге», Санкт-Петербург, 2001-2007; ХIХ Российском съезде физиологического общества им. И.П. Павлова с международным участием, Екатеринбург, 2004; VI Международной конференции «Проблемы загрязнения окружающей среды», Пермь-Казань, 2005; III съезде Российского научного общества иммунологов, Екатеринбург, 2004; I-V конференциях иммунологов Урала, Екатеринбург, 2001; Пермь, 2002; Челябинск, 2003; Уфа, 2005; Оренбург, 2006; I-II конференциях молодых учёных «Современные проблемы микробиологии, иммунологии и экологии», Пермь, 1999, 2002.
Публикации. Материалы диссертационной работы обобщены в 46 печатных работах, в том числе 15 экспериментальных статьях и 31 материалах конференций.
Объем и структура работы. Диссертация изложена на 251 странице, содержит 35 таблиц, 49 рисунков и состоит из введения, литературного обзора, описания объектов и методов исследования, 5 глав результатов собственных исследований, обсуждения, выводов, списка цитируемой литературы, включающего 448 наименований, в том числе 124 на русском и 324 на английском языках.
Место проведения работы. Работа является частью исследований, выполняемых в аналитической лаборатории ИЭГМ УрО РАН (зав. – к.г.-м.н. М.А. Шишкин) совместно с лабораторией экологической иммунологии (зав. – к.м.н. Б.А. Бахметьев) по изучению механизмов иммуномодулирующих эффектов гормонов, продукция которых изменяется на фоне экологического воздействия. Исследования по проблеме нейроэндокринной регуляции иммуногенеза были инициированы профессором, заслуженным деятелем науки РФ Н.Н. Кеворковым. Научные положения диссертации и выводы, вытекающие из анализа полученного экспериментального материала, базируются на результатах собственных исследований автора.
Автор выражает искреннюю благодарность М.А. Шишкину, к.х.н. С.П. Тендряковой, профессору М.В. Черешневой, за внимание и моральную поддержку. Автор особо признателен сотрудникам группы радиоизотопных исследований к.б.н. Т.А. Баевой, инженеру Е.Г. Чижовой, магистрантам кафедры микробиологии и иммунологии Пермского государственного университета К.Г. Горшковой и И.Л. Шаравьёвой, способствующим завершению настоящей работы и чей вклад в определённые разделы исследований отражён в приведённых в списке литературы публикациях. Автор благодарит главного специалиста Муниципального управления здравоохранением Ростехнадзора, к.м.н. В.Г. Рыжаенкова за помощь в проведении иммуноферментного анализа.
Глубокую благодарность и признательность автор выражает своим учителям и наставникам академику РАН и РАМН В.А. Черешневу и доценту Ю.И. Шилову, оказавшим большое влияние на выбор целей научного поиска и формирование научного мировоззрения автора.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы исследования. В работе использовали белых беспородных мышей массой 20-22 г и лейкоциты периферической венозной крови, полученной от здоровых людей – добровольцев мужского пола в возрасте 19-35 лет.
Для экспериментального моделирования реакции стресс использовали ротационную модель. Ротация мышей производилась в течение 60 мин по 10 мин с перерывами по 5 мин при 78 об/мин. Роль опиатных рецепторов в постстрессорных изменениях иммунных реакций исследовали путем их блокады налоксоном гидрохлоридом и налтриндолом гидрохлоридом. Налоксона гидрохлорид (DuPont, США) в разовой дозе 0,2 мг/кг массы тела и селективный антагонист d-опиатных рецепторов налтриндола гидрохлорид (ICN, США) в дозе 0,1 мг/кг вводили животным подкожно однократно за 20 мин до ротации (Ашмарин, 1988; Михайлова и др., 1992; Croock et al., 1992). В дальнейших экспериментах в системе in vivo дозы опиатных антогонистов не изменялись. Иммунизацию животных производили через 1 ч после окончания ротации.
При исследовании иммунорегуляторных эффектов опиоидных пептидов in vivo β-эндорфин (Sigma, США) в диапазоне доз от 100 мкг/кг до 0,0005 мкг/кг вводили однократно внутрибрюшинно в объеме 0,2 мл. Контролем для животных, получавших β-эндорфин, служили мыши, которым вводили по той же схеме 0,9% NaCl. μ-агонист DAGO ([d-Ala2,N-Me-Phe4,Gly5-ol]-энкефалин и δ-агонист DADLE ([d–Ala2,d-Leu5] - энкефалин) (Sigma, США) в диапазоне 10 – 0,0001 мкг/кг вводили по схеме аналогичной введению β-эндорфина. Иммунизацию животных производили через 1 ч после введения опиоидных пептидов.
Гидрокортизона ацетат (Гедеон Рихтер, Венгрия) в дозе 50 мг/кг массы тела вводили однократно внутрибрюшинно. Адреналина гидрохлорид (Московский эндокринный завод, Россия) вводили подкожно однократно в дозе 1 мг/кг. Налоксон и селективный антагонист d-опиатных рецепторов налтриндол вводили подкожно за 20 мин до введения гормонов (3 инъекции через 2,5 ч в группе с гидрокортизоном и 1 инъекция в группе с адреналином). Контролем служили интактные мыши, подвергшиеся иммунизации, но не получавшие препаратов. Дополнительным контролем для животных, получавших гидрокортизон и опиоидные пептиды, служили мыши, получавшие по той же схеме изотонический раствор хлорида натрия. Иммунизацию опытных и контрольных мышей проводили одномоментно через 3 ч от начала эксперимента в группах с гидрокортизоном, через 30 мин - в группах с адреналином.
Для моделирования локального иммунного ответа животных иммунизировали эритроцитами барана (108 клеток вводили подкожно в подошвенную поверхность правой стопы). На 4-е сутки вводили разрешающую дозу антигена (108 клеток). На 5-е сутки оценивали выраженность иммунного воспаления при реакции ГЗТ путём регистрации толщины (инженерным микрометром) и массы (на торсионных весах) опытной и контрольной стопы; количество ядросодержащих клеток (ЯСК); интенсивность антителогенеза методом локального гемолиза в геле агарозы (Jerne, Nordin, 1963). Оценку фагоцитарной активности клеток периферической крови, селезенки, регионарного и отдаленного подколенных лимфатических узлов проводили методом В.Н. Каплина с соавт. (Каплин, 1992, 1996) в модификации (Шилов и др., 1997, 1998).
Нефракционированную клеточную взвесь получали путём отстаивания верхнего слоя плазмы крови с лейкоцитами. Выделение фракции мононуклеаров и нейтрофилов проводили на градиенте плотности фиколл-верографин. Разделение моноцитов и лимфоцитов проводили методом адгезии на чашках Петри. CD4+ Т-клетки выделяли при помощи набора магнитных бус Dynabeads M-450 CD4 (Invitrogen, США). Культивирование клеток проводили в течение 24, 48 и 72 ч в пластиковых 24 и 96-луночных планшетах (Orange Scientific, Бельгия) в соответствии с традиционными методиками с использованием полной питательной среды, приготовленной на основе RPMI 1640 или среды 199 (Биолот, Россия) с добавлением 10 mM HEPES, 2 mM L-глутамина (Sigma, США), 100 мкг/мл гентамицина и 10% эмбриональной телячьей сыворотки (Биолот, Россия) или аутоплазмы во влажной атмосфере с 5% СО2 при 370С.
Пролиферативную активность оценивали по включению 3H-метилтимидина. Радиоактивность проб определяли на жидкостном сцинтилляционном счетчике Guardian (Wallac, Финляндия). Для определения концентрации IL-1β, TNF-a, IL-6, IL-8, IL-1ra, IL-2, IL-4 и IFN-γ в супернатантах культур клеток использовали спектрофотометр Униплан (Пикон, Россия) и иммуноферментные тест-системы производства ООО Протеиновый контур, ООО Цитокин, Санкт-Петербург, Вектор-Бест, Новосибирск. В экспериментах in vitro использовали агонист δ,μ-опиатных рецепторов β-эндорфин в концентрациях 10-7-10-12М; меланотропин потенцирующий фактор (MPF) - фрагмент 88-91 β-липотропина (Lys-Lys-Gly-Glu) в концентрациях 10-7-10-12М; μ-агонист опиатных рецепторов DAGO ([d-Ala2,N-Me-Phe4,Gly5-ol]-энкефалин) в концентрациях 10-7 10-12М; δ-агонист опиатных рецепторов DADLE ([d–Ala2,d-Leu5]-энкефалин) в концентрациях 10-7–10-12М; неселективный антагонист опиатных рецепторов налоксона гидрохлорид и селективный антагонист δ-рецепторов налтриндола гидрохлорид в концентрациях 10-6, 10-8, 10-10М; липополисахарид (ЛПС) Escherichia coli O26:B6 - 0,1 мкг/мл (Sigma, США), фитогемагглютинин (ФГА) – 1,25; 2,5; 5,0; 10,0; 20,0 мкг/мл (Sigma, США), диклофенак натрия (ДН) 25 мкг/мл, моноклональные анти-IL-1b антитела – 2 мкг/мл.
Полученные данные обрабатывали с помощью многофакторного дисперсионного анализа для парных данных и корреляционного анализа. Достоверность различий между группами оценивали с помощью t-критерия Стьюдента и критерия Фишера наименьшей значимой разницы. Сортировку и обработку данных проводили на компьютере IBM PC c использованием программ Statistica for Windows 6.0 (Statsoft, Inc., США) и DIASTA (Московский государственный университет, Россия).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ
Влияние ротационного стресса на показатели иммунитета. Роль опиатных рецепторов. В большинстве опубликованных работ, посвященных изучению влияния стресса на иммунный ответ, исследуются изменения системного иммунного ответа в условиях внутривенной или внутрибрюшинной иммунизации. Принимая во внимание разные компоненты внутрисистемной регуляции общих и локальных форм иммунного ответа, представлялось целесообразным исследование эффектов стресса и блокады опиатных рецепторов в условиях развития локальной формы иммунного ответа.
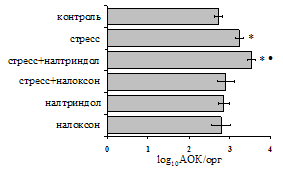
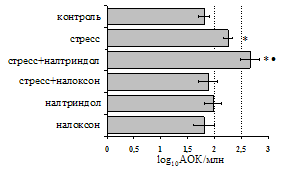
Как видно из рис. 1, в индуктивную фазу иммунного ответа на фоне стресса наблюдается увеличение числа АОК в лимфатическом узле (ЛУ) и усиление степени выраженности реакции ГЗТ. Блокада δ-опиатных рецепторов приводит к ещё более выраженной активации антителогенеза, в то время как
А Б
В Г
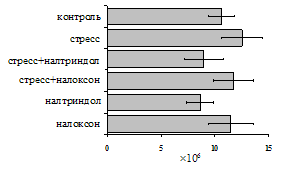
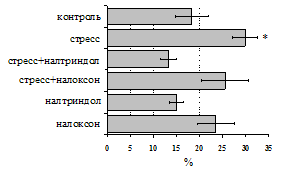
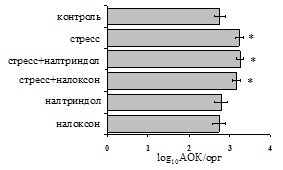
Рис. 1. Влияние ротационного стресса в условиях блокады опиатных рецепторов на абсолютное (А) и относительное (Б) число АОК, количество ЯСК в регионарном лимфатическом узле (В) и выраженность реакции ГЗТ (Г) в индуктивную фазу иммунного ответа. Здесь и на рис. 2: * - p<0,05 к контролю; - p<0,05 к стрессу.
на фоне налоксона стимулирующий эффект ротационного стресса на антителогенез отменяется. Стрессиндуцированное усиление выраженности реакции ГЗТ отменяется как налоксоном, так и налтриндолом. Изолированное введение животным налтриндола или налоксона на количество АОК и степень выраженности ГЗТ влияния не оказывает. В эффекторную фазу иммунного ответа ротационный стресс (рис. 2) стимулирует как клеточный, так и гуморальный ответ, однако, в отличие от индуктивной фазы, на фоне блокады опиатных рецепторов эффекты стресса не модифицируются. Таким образом, стимуляция опиатных рецепторов в индуктивную фазу иммунного ответа играет важную роль в стрессиндуцированных изменениях иммуногенеза и ответственна за активацию функций иммунной системы при стрессе.
Влияние гидрокортизона и адреналина на локальный иммунный ответ в условиях блокады опиатных рецепторов. В процессе развития стрессреакции основные стрессреализующие факторы глюкокортикоиды,
А Б
В Г
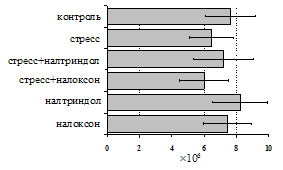
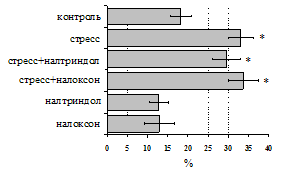
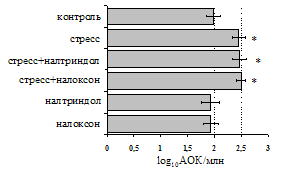
Рис. 2. Влияние ротационного стресса в условиях блокады опиатных рецепторов на абсолютное (А) и относительное (Б) число АОК, количество ЯСК в регионарном лимфатическом узле (В) и выраженность реакции ГЗТ (Г) в эффекторную фазу иммунного ответа.
Таблица 1. Влияние гидрокортизона в условиях блокады m- и d-опиатных рецепторов на число АОК, количество ЯСК в лимфатическом узле и выраженность реакции ГЗТ в индуктивную фазу иммунного ответа
| Экспериментальное воздействие |
Лимфатический узел |
||
ЯСК на орган(´106) |
log10 АОК на 106 ЯСК |
log10 АОК на орган |
|
| Интактные животные (контроль), n=19 | 8,93±1,35 | 2,15±0,15 | 3,01±0,17 |
| (142) | (1014) | ||
| Физиологический | 7,79±1,08 | 2,27±0,16 | 3,10±0,13 |
| раствор, n=16 | (186) | (1252) | |
| Гидрокортизон, n=19 | 4,98±0,68*# | 1,64±0,23 | 2,17±0,25*# |
| (43) | (149) | ||
| Гидрокортизон | 6,71±0,75a | 0,96±0,21*#a | 1,50±0,31*# |
| +Налоксон, n=15 | (9) | (32) | |
| Гидрокортизон | 5,87±0,71 | 0,55±0,14*#a | 1,07±0,22*#a |
| +налтриндол, n=17 | (4) | (12) | |
| Налоксон, n=10 | 7,76±1,27 | 2,24±0,08 | 3,07±0,14 |
| (173) | (1187) | ||
| Налтриндол, n=12 | 8,38±1,37 | 1,93±0,11 | 2,79±0,11 |
| (86) | (622) | ||
Примечание. Здесь и в табл. 2, 3, 4 в скобках указана средняя геометрическая числа АОК (антилогарифм из средней арифметической log10 числа АОК). * - p<0,05 к контролю; # - p<0,05 к физиологическому раствору; а - p<0,05 к гидрокортизону по непарному t-критерию Стъюдента..
катехоламины и эндогенные опиоиды находятся в тесной взаимосвязи и оказывают друг на друга взаимное регуляторное влияние (O`Connor, 2000). Как видно из табл. 1, в индуктивную фазу иммунного ответа гидрокортизон снижает количество ядросодержащих клеток в лимфатическом узле и абсолютное число АОК. При введении гидрокортизона на фоне блокады опиатных рецепторов налоксоном и налтриндолом регистрируется отмена индуцированного гидрокортизоном снижения количества ЯСК и ещё более выраженное угнетение антителогенеза по абсолютным и относительным показателям,
Таблица 2. Влияние адреналина в условиях блокады m- и d-опиатных рецепторов на число АОК, количество ЯСК в лимфатическом узле и выраженность реакции ГЗТ в индуктивную фазу иммунного ответа
| Экспериментальное воздействие |
Лимфатический узел |
|||
ЯСК наорган (´106) |
log10 АОК на 106 ЯСК |
log10 АОК на орган |
||
|
Интактные животные (контроль) n=11 |
6,29±1,04 | 2,43±0,16 | 3,15±0,22 | |
| (268) | (1403) | |||
| Адреналин, n=9 | 5,71±0,74 | 2,07±0,14 | 2,79±0,16 | |
| (118) | (618) | |||
Адреналин |
7,37±2,08 | 1,94±0,15* | 2,73±0,24 | |
| +Налоксон, n=6 | (86) | (541) | ||
| Адреналин | 6,62±1,30 | 1,78±0,25* | 2,45±0,32 | |
| +налтриндол, n=11 | (60) | (283) | ||
| Налоксон, n=10 | 7,76±1,27 | 2,24±0,08 | 3,07±0,14 | |
| (173) | (1187) | |||
| Налтриндол, n=9 | 8,07±1,82 | 2,08±1,97 | 2,90±0,13 | |
| (121) | (803) | |||
особенно ярко проявляющееся в условиях блокады d-рецепторов. Изолированное введение экспериментальным животным налтриндола и налоксона на исследуемые показатели влияния не оказывает. В эффекторную фазу иммунного ответа гидрокортизон угнетает количество ядросодержащих клеток в лимфатическом узле, абсолютное число АОК, однако в отличие от индуктивного периода на фоне блокады опиатных рецепторов эффекты гидрокортизона на показатели клеточности и антителогенеза не модифицируются.
Несколько иная картина наблюдается при анализе эффектов адреналина. Как видно из табл. 2, в индуктивную фазу адреналин оказывает статистически достоверный эффект на антителогенез по относительным показателям (F=5,28; p<0,03), несмотря на то, что при межгрупповом сравнении по отношению к контролю угнетение относительного количества АОК имеет место только при комбинации адреналина с опиатными
Таблица 3. Влияние β-эндорфина на число АОК, клеточность и выраженность реакции ГЗТ в регионарном лимфатическом узле
| Экспериментальное воздействие (β-эндорфин) | Число живот-ных | Лимфатический узел | Интенсив-ность ГЗТ | ||
|
ЯСК на орган (*106) |
Log10 АОК на 106 ЯСК |
Log10 АОК на Орган |
ИР по массе стопы, % · | ||
| Контроль | 9 | 4,62±0,89 | 2,43±0,10 | 3,04±0,01 | 17,22±2,41 |
| (271,28) | (1088,94) | ||||
| 100 мкг/кг | 9 | 4,33±0,71 | 1,98±0,22* | 2,57±0,26* | 19,84±3,68 |
| (95,28) | (370,54) | ||||
| 10 мкг/кг | 8 | 5,25±0,70 | 2,06±0,26 | 2,76±0,25 | 23,92±3,46 |
| (115,56) | (569,35) | ||||
| 1 мкг/кг | 9 | 4,87±1,06 | 2,46±0,18 | 3,05±0,16 | 20,25±4,25 |
| (286,09) | (1128,59) | ||||
| 0,1 мкг/кг | 8 | 7,55±1,35 | 2,25±0,18 | 3,08±0,13 | 18,82±6,17 |
| (117,27) | (1198,91) | ||||
| 0,01 мкг/кг | 9 | 5,33±1,13 | 2,68±0,09 | 3,34±0,13 | 26,72±3,88 |
| (474,99) | (2192,95) | ||||
| 0,001 мкг/кг | 8 | 5,90±1,73 | 2,62±0,73 | 3,21±0,13 | 21,27±4,88 |
| (421,49) | (1607,54) | ||||
| 0,0005 мкг/кг | 9 | 5,82±0,81 | 2,87±0,08* | 3,61±0,07* | 28,72±4,10 |
| (743,72) | (4031,74) | ||||
Примечание. Здесь и в табл. 4: · - индекс реакции (ИР): И.Р. = (Ро–Рк)/Рк·100%, где Ро и Рк - показатели массы в опытной и контрольной конечностях. * - p<0,05 к контролю по t-критерию Фишера наименьшей значимой разницы.
антагонистами. В эффекторную фазу иммунного ответа адреналин на антителогенез в лимфатическом узле не влияет. Таким образом, блокада опиатных рецепторов в индуктивную фазу иммунного ответа приводит к существенной модификации иммунорегуляторных эффектов глюкокортикоидов и катехоламинов, связанной с изменениями секреции β-эндорфина в ответ на введение гидрокортизона или адреналина (Mougey et al., 1986; Bagdy et al., 1989; Goodwin et al., 1992).
Влияние β-эндорфина на показатели клеточного и гуморального иммунитета. Степень выраженности эффектов β-эндорфина в системе in vivo
Таблица 4. Влияние β-эндорфина в условиях блокады опиатных рецепторов на число АОК, клеточность и выраженность реакции ГЗТ в регионарном лимфатическом узле в индуктивную фазу иммунного ответа
| Экспериментальное воздействие | Чис-ло жи-во-тных | Лимфатический узел | Интенсив-ность ГЗТ | ||
|
ЯСК на орган (´106) |
log10 АОК на 106 ЯСК |
log10 АОК на орган |
ИР по массе стопы, % | ||
| Контроль | 18 | 5,34±0,62 | 2,35±0,10 | 3,02±0,11 | 22,43±3,19 |
| (223,55) | (1043,31) | ||||
| β-эндорфин | 18 | 4,71±0,51 | 1,99±0,14* | 2,36±0,15* | 22,16±2,73 |
| (100 мкг/кг) | (98,80) | (423,03) | |||
| β-эндорфин | 17 | 5,98±0,66 | 2,67±0,07* | 3,40±0,08* | 27,95±2,73 |
| (0,0005 мкг/кг) | (465,21) | (2519,33) | |||
| β-эндорфин | 11 | 5,64±0,97 | 2,21±0,14 | 2,87±0,18 | 23,79±4,27 |
| (100 мкг/кг) + | (161,61) | (747,62) | |||
| налоксон | |||||
| β-эндорфин | 12 | 5,95±0,66 | 2,16±0,12 | 2,90±0,14 | 28,57±6,77 |
| (0,0005 мкг/кг) + | (143,12) | (798,77) | |||
| налоксон | |||||
| β-эндорфин | 11 | 7,18±41,24 | 2,50±0,07 | 3,29±0,05* | 23,32±4,17 |
| (100 мкг/кг) + | (314,71) | (1945,42) | |||
| налтриндол | |||||
| β-эндорфин | 11 | 9,07±1,24* | 2,52±0,09 | 3,43±0,12* | 25,67±3,12 |
| (0,0005 мкг/кг) + | (327,67) | (2673,98) | |||
| налтриндол | |||||
| Налоксон | 12 | 6,40±0,73 | 2,23±0,09 | 3,01±0,11 | 16,08±2,02 |
| (170,40) | (1013,98) | ||||
| Налтриндол | 8 | 6,98±0,55 | 2,04±0,24 | 2,87±0,21 | 18,36±1,92 |
| (110,87) | (755,38) | ||||
Примечание. * - p<0,05 к контролю по непарному t-критерию Стьюдента.
напрямую зависит от вводимой дозы пептида (табл. 3). Пептид оказывает разнонаправленный эффект на гуморальный иммунный ответ, угнетающий в дозе 100 мкг/кг и стимулирующий в дозе 0,0005 мкг/кг образование АОК в регионарном ЛУ. При этом статистически достоверного влияния β-эндорфина на клеточность ЛУ и степень выраженности реакции ГЗТ не обнаруживается. Таким образом, β-эндорфин в системе in vivo в зависимости от дозы как усиливает, так и угнетает образование антителопродуцентов.
Данные о влиянии β-эндорфина на фоне блокады опиатных рецепторов представлены в табл. 4. Блокада рецепторов неселективным антагонистом налоксоном отменяет как угнетающий эффект дозы 100 мкг/кг, так и стимулирующий эффект дозы 0,0005 мкг/кг на относительное и абсолютное количество АОК. В то же время введение мышам β-эндорфина на фоне блокады δ-рецепторов налтриндолом по абсолютным показателям не отменяет стимулирующего эффекта низкой (0,0005 мкг/кг) дозы пептида и приводит к увеличение числа АОК в ответ на введение животным высокой дозы (100 мкг/кг). Кроме этого, введение пептида в дозе 0,0005 мкг/кг на фоне налтриндола приводит к статистически достоверному увеличению клеточности ЛУ по сравнению с контролем. На степень выраженности иммунного воспаления комбинация β-эндорфина с антагонистами опиатных рецепторов влияния не оказывает. Таким образом, β-эндорфин в зависимости от дозы оказывает разнонаправленное влияние на образование антителопродуцентов, не влияя
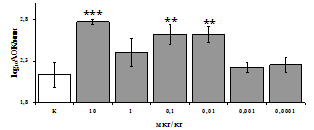
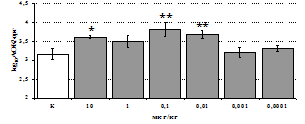
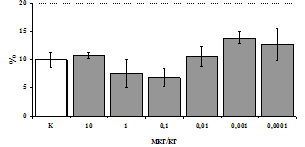
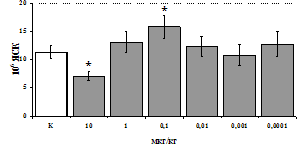
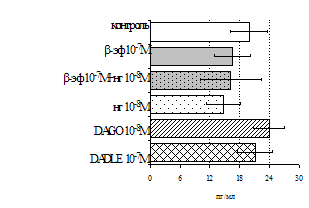
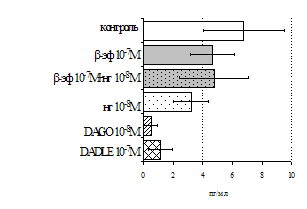
Рис. 3. Влияние DADLE на относительное и абсолютное число АОК, выраженность реакции ГЗТ и клеточность в регионарном лимфатическом узле в индуктивную фазу иммунного ответа.
* - p<0,05; ** - p<0,01; *** - p<0,001 к контролю по t-критерию Фишера наименьшей значимой разницы.
на клеточноопосредованный ответ. Способность пептида взаимодействовать с d-рецептором проявилась только при введении высокой дозы, что, в свою, очередь указывает на возможность реализации через d-рецептор иммуносупрессивных эффектов, напротив блокада m-рецепторов отменяет эффекты пептида независимо от вводимой дозы. В эффекторную фазу влияния β-эндорфина на иммунный ответ не выявляется.
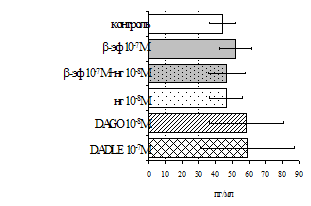
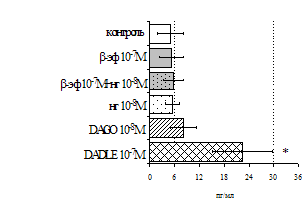
Параллельно нами изучалось сравнительное влияние селективных m и d-агонистов на выраженность локального иммунного ответа. Как видно из рис. 3, в индуктивную фазу иммунного ответа введение d-агониста DADLE в дозах 10,0; 0,1; 0,01 мкг/мл стимулирует количество АОК по абсолютным и относительным параметрам, не влияет на степень выраженности иммунного воспаления (ГЗТ) и оказывает разнонаправленное действие на клеточность регионарного лимфатического узла, при этом в дозе 10 мкг/кг угнетая, а в дозе 0,1 мкг/кг - увеличивая содержание ЯСК. Введение m-агониста DAGO статистически достоверно влияет только на относительное число АОК и клеточность регионарного лимфатического узла. В дозах 10,0; 1,0; 0,0001 мкг/кг пептид активирует образование антителопродуцентов по относительным показателям и в диапазоне доз 10-
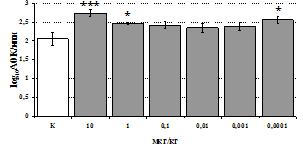
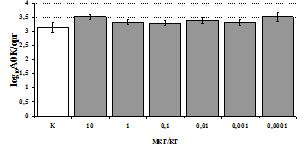
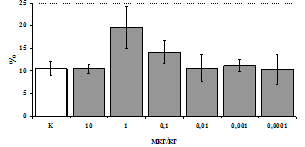
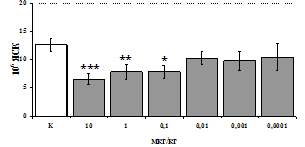
Рис. 4. Влияние DAGO на относительное и абсолютное число АОК, выраженность реакции ГЗТ и клеточность в регионарном лимфатическом узле в индуктивную фазу иммунного ответа.
* - p<0,05; ** - p<0,01; *** - p<0,001 к контролю по t-критерию Фишера наименьшей значимой разницы.
0.1 мкг/кг угнетает количество ЯСК (рис. 4). Таким образом, по нашим данным, в системе in vivo эффекты b-эндорфина и аналогов энкефалинов с m,d-селективным спектром связывания DAGO и DADLE значительно варьируют по направленности действия, эффективному диапазону доз, взаимодействию с опиатных рецепторов различных типов, а так же зависят от этапа, на котором конкретный опиоидный пептид вмешивается в развитие иммунных реакций. В то же время наиболее выраженное активирующее влияние наблюдается при введении экспериментальным животным селективного агониста d-рецепторов DADLE.
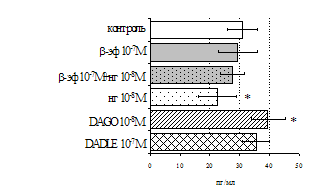
Влияние β-эндорфина, 88-91 фрагмента липотропина MPF, селективных лигангдов DAGO, DADLE на пролиферативный ответ лимфоцитов. Степень выраженности эффектов исследуемых опиоидных пептидов в системе in vitro зависит от их концентрации и присутствия митогена в культуральной среде. Все лиганды опиатных рецепторов проявляют активность только на стимулированных митогеном культурах. Как видно из рис. 5, b-эндорфин в концентрации 10-7М статистически значимо усиливает пролиферативный ответ лимфоцитов в культурах с ФГА 5 мкг/мл. Внесение пептида в культуры
A 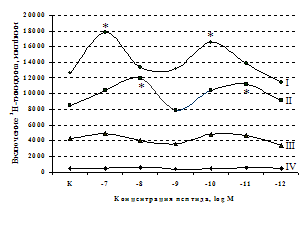
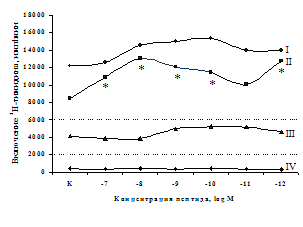
Рис. 5. Влияние b-эндорфина (A) и DAGO (Б) на ФГА-индуцированный пролиферативный ответ лимфоцитов.
Здесь и на рис. 6: I - ФГА 5 мкг/мл, II - ФГА 2,5 мкг/мл, III - ФГА 1,25 мкг/мл, IV без внесения ФГА. Число наблюдений в группах с b-эндорфином n=9, в группах с DAGO - n=8. * - р<0.05 к контролю по парному t-критерию Фишера наименьшей значимой разницы..
в концентрации 10-8М приводит к стимуляции реакции бласттрансформации в присутствии ФГА 2,5 мкг/мл. Низкие (10-10, 10-11М) концентрации b-эндорфина стимулируют пролиферативный ответ в культурах с ФГА 5,0 и 2,5 мкг/мл соответственно. Внесение b-эндорфина в концентрации 10-12М, отражающей фоновый уровень пептида в плазме крови, не оказывает существенного влияния на пролиферацию лимфоцитов. А.А. Зозулей и С.Ф. Пшеничкиным (1990) высказано предположение, что иммуномодулирующие эффекты b-эндорфина могут проявляться через С-концевой участок пептидной цепи, невзаимодействующий с d-, m-рецепторами. В связи с этим представлял интерес анализ эффектов С-концевого тетрапептида b-эндорфина MPF (меланотропин-потенциирующего фактора) на пролиферативную активность лимфоцитов периферической крови. Как видно из рис. 6, MPF ни в одной из исследуемых концентраций статистически значимых эффектов на спонтанную и индуцированную митогеном пролиферацию лимфоцитов не оказывает. Это позволяет предположить, что выявленный стимулирующий эффект b-эндорфина на пролиферативную активность лимфоцитов не опосредуется через его С-концевой участок.
В связи с тем, что b-эндорфин N-концевой последовательностью связывается как с m-, так и d-опиатными рецепторами и не ясно, какой тип рецепторов в данном случае является основным проводником сигнала с поверхности клетки, мы сопоставили его эффекты на
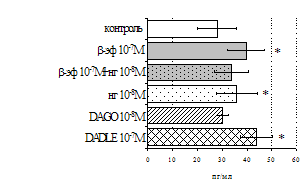
A 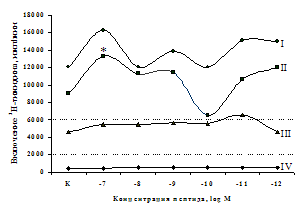
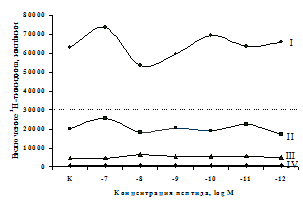
Рис. 6. Влияние DADLE (А) и MPF (Б) на ФГА-индуцированный пролиферативный ответ лимфоцитов.
Число наблюдений n=8. пролиферацию с эффектами селективных m- и d-агонистов. Данные о влиянии синтетического селективного агониста μ-опиатных рецепторов DAGO на пролиферативную активность лимфоцитов периферической крови представлены в рис. 5. Анализ зависимости эффектов от концентрации показал, что DAGO в высоких и низких концентрациях достоверно усиливает пролиферативный ответ лимфоцитов исключительно в присутствии субоптимальной концентрации (2,5 мкг/мл) митогена. На спонтанный пролиферативный ответ DAGO подобно b-эндорфину влияния не оказывает.
Аналогичные результаты получены при анализе влияния селективного d-агониста DADLE на пролиферацию лимфоцитов (см. рис. 6). Выявлено, что DADLE усиливает включение 3Н-тимидина лимфоцитами по сравнению с контролем в присутствии ФГА 2,5 мкг/мл и высокой концентрации (10-7 М) данного пептида.
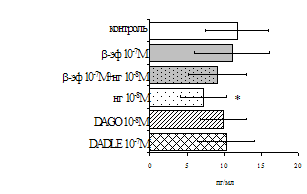
В дальнейшем проводилась сравнительная оценка влияния β-эндорфина на пролиферативный ответ на фоне блокады опиатных рецепторов в нефракционированных и фракционированных, очищенных от моноцитов, лимфоцитарных культурах. Как видно из рис. 7, в нефракционированной лейкоцитарной суспензии стимулирующий эффект b-эндорфина на пролиферативный ответ не отменяется, а напротив, усиливается. При удалении моноцитов из фракции мононуклеаров b-эндорфин (F=13,07; p=0,006) и налоксон (F=10,21; p=0,011) оказывают высоко достоверные
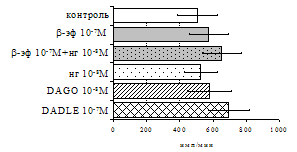
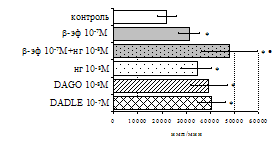
Рис. 7. Эффекты b-эндорфина на фоне блокады опиатных рецепторов, DAGO и DADLE на спонтанную (A) и индуцированную ФГА 2,5 мкг/мл (Б) пролиферативную активность лимфоцитов в нефракционированных клеточных культурах.
* - р<0,05 к контролю по парному t-критерию Стъюдента.
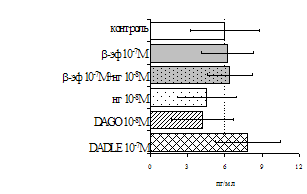
самостоятельные эффекты на ФГА-индуцированную пролиферацию, не проявляя статистически значимого взаимодействия между собой (F=4,08; p=0,074). Последующее сравнение средних величин показало, что направленность эффектов опиоидных пептидов противоположна влиянию, оказываемому b-эндорфином и налоксоном в присутствии фракции моноцитов (рис. 8). В частности, выраженное угнетение пролиферации лимфоцитов как по сравнению с контролем, так и с b-эндорфином наблюдается при совместном внесении в культуры b-эндорфина и налоксона. Угнетающий эффект на захват 3Н-тимидина лимфоцитами оказывает селективный m-агонист DAGO. Селективный агонист d-рецепторов DADLE на ФГА-индуцированную пролиферативную активность лимфоцитарной фракции не влияет. При этом эффекты исследуемых опиоидов в аналогичных культурах без митогена не обнаруживаются. Проведенный корреляционный анализ выявил статистически достоверную (r=0,73; р<0,05) зависимость между интенсивностью пролиферации лимфоцитов в нефракционированных культурах под воздействием b-эндорфина и в аналогичных культурах, очищенных от фракции моноцитов.
Анализ роли d-рецепторов в регуляции пролиферативного ответа в нефракционированной лейкоцитарной суспензии и фракции лимфоцитов (рис. 9) показал отмену стимулирующего эффекта b-эндорфина налтриндолом во фракции лейкоцитов. Во фракции лимфоцитов b-эндорфин и налтриндол на степень выраженности пролиферации не влияют, что свидетельствует о возможной
А Б
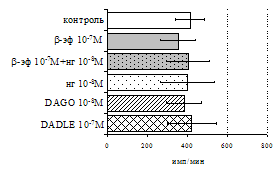
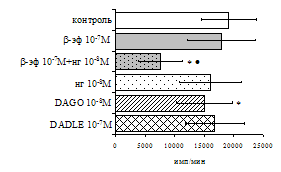
Рис. 8. Эффекты b-эндорфина на фоне блокады опиатных рецепторов, DAGO и DADLE на спонтанную (A) и индуцированную ФГА 2,5 мкг/мл (Б) пролиферативную активность лимфоцитов в клеточных культурах, очищенных от фракции моноцитов.
* - р<0,05 к контролю по парному t-критерию Стъюдента.
А Б
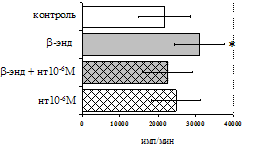
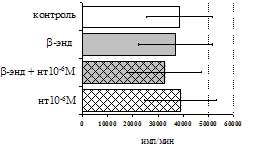
Рис. 9. Эффекты b-эндорфина на фоне блокады d-опиатных рецепторов на индуцированную ФГА 2,5 мкг/мл пролиферативную активность лимфоцитов во фракции лейкоцитов (А) и фракции лимфоцитов (Б).
* - р<0,05 к контролю по парному t-критерию Стъюдента.
реализации стимулирующего эффекта пептида через d-рецептор, но только в присутствии моноцитов.
Учитывая важную регуляторную роль моноцитов, в дальнейшем мы попытались оценить роль IL-1b и продуктов циклооксигеназного цикла в b-эндорфинопосредованной регуляции пролиферативного ответа лимфоцитов в присутствии ФГА (рис. 10А). Все обследованные здоровые доноры были разделены по индивидуальной чувствительности к b-эндорфину на две группы: у 1-й группы пептид стимулировал
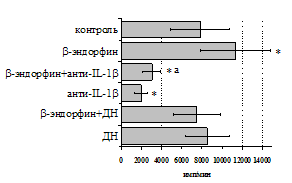
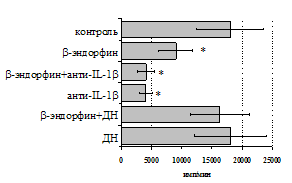
Рис. 10. Влияние b-эндорфина на ФГА-индуцированный пролиферативный ответ лимфоцитов в присутствии анти-IL-1b антител и на фоне блокады синтеза простагландинов ДН у доноров 1-й (А, n=9) и 2-й (Б, n=11) групп.
* - р<0,05 к контролю; а - р< 0,05 к анти-IL-1b по парному t-критерию Стъюдента.
пролиферативный ответ, а у 2-й – угнетал. В первой группе доноров на фоне моноклональных антител к IL-1b наблюдается резкое снижение пролиферативной активности, в то же время при внесении в культуры анти-IL-1b-антител в присутствии b-эндорфина наблюдается некоторое усиление пролиферативного ответа, достоверно отличающееся от культур с анти-IL-1b-антителами, но по сравнению с контролем уровень захвата метки был так же достоверно ниже. Как видно рис. 10Б, у второй группы доноров на фоне анти-IL-1b-антител интенсивность пролиферативного ответа в присутствии b-эндорфина не изменяется.
А
без ФГА ФГА 2,5 мкг/мл
Б
без ФГА ФГА 2,5 мкг/мл
Рис. 11. Влияние b-эндорфина, DAGO, DADLE на продукцию IFN-g в нефракционированной (А) и фракционированной культурах (Б).
Здесь и на рис. 12 число наблюдений n=10. * - р<0,05 к контролю по парному t-критерию Стъюдента.
А
без ФГА ФГА 2,5 мкг/мл
Б
без ФГА ФГА 2,5 мкг/мл
Рис. 12. Влияние b-эндорфина, DAGO, DADLE на продукцию IL-4 в нефракционированной (А) и фракционированной клеточных культурах (Б).
* - р<0,05 к контролю по парному t-критерию Стъюдента.
При культивировании лейкоцитов в присутствии диклофенака натрия как стимулирующий, так и угнетающий эффект b-эндорфина на пролиферативный ответ нивелируется, что подтверждает данные о возможном участии простагландинов (простагландина E2 (PGЕ2), в частности) в регуляции функциональной активности лимфоцитов под воздействием опиоидных пептидов. Таким образом, регуляция функциональной активности лимфоцитов b-эндорфином может опосредоваться как системой IL-1, так и простагландинами.
Влияние β-эндорфина и селективных лигандов опиатных рецепторов на процессы клеточной кооперации и переключение Th1/Th2 цитокинового профиля. Следующим этапом исследований являлось изучение роли b-эндорфина в регуляции продукции IL-4 и IFN-g в супернатантах нефракционированной лейкоцитарной взвеси, лимфоцитарной фракции и культуре CD4+ клеток. Как показано на рис. 11, уровень g-IFN в супернатантах под воздействием b-эндорфина, а также в случае комбинации b-эндорфина с налоксоном не отличается от контроля как в нефракционированных культурах, так и в культурах, очищенных от моноцитов, независимо от присутствия ФГА в среде культивирования. Однако в фракционированных клеточных культурах, стимулированных митогеном, регистрируется эффект селективного d-агониста DADLE на продукцию IFN-g, что, очевидно, обусловлено его прямым эффектом на рецепторные структуры клеточной поверхности лимфоцитов.
На рис. 12 приведены результаты исследования влияния b-эндорфина, DAGO, DADLE на продукцию IL-4 в нефракционированной лейкацитарной суспензии и фракции лимфоцитов. По нашим данным, под воздействием налоксона и DAGO в культурах без митогена регистрируются угнетающий и стимулирующий эффекты соответственно. Выраженный стимулирующий эффект на ФГА-индуцированную продукцию IL-4 оказывают b-эндорфин, налоксон и DADLE. В очищенной фракции лимфоцитов значимых эффектов исследуемые соединения на продукцию IL-4 не выявляется, за исключением угнетающего эффекта
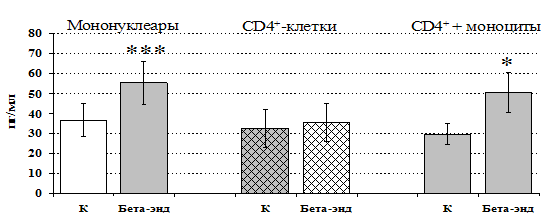
Рис 13. Влияние b-эндорфина 10-7 М на продукцию IL-4 фракцией мононуклеаров, CD4+-лимфоцитами и CD4+-лимфоцитами в присутствии моноцитов в присутствии ФГА 2,5 мкг/мл.
* - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Стъюдента.
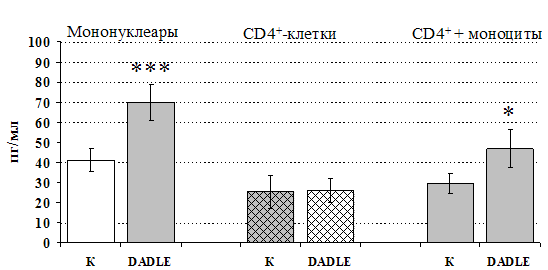
Рис 14. Влияние DADLE 10-7 М на продукцию IL-4 фракцией мононуклеаров, CD4+-лимфоцитами и CD4+-лимфоцитами в присутствии моноцитов в присутствии ФГА 2,5 мкг/мл.
* - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Стъюдента.
налоксона, зарегистрированного в культурах без добавления митогена. Корреляционный анализ выявил отрицательную зависимость (r=-0,68; р<0,05) между интенсивностью пролиферации и уровнем IL-4 в культурах с совместным внесением b-эндорфина и налоксона в присутствии ФГА. Таким образом, b-эндорфин, налоксон и селективный агонист δ-рецепторов DADLE, усиливая пролиферацию, способствуют изменению соотношения Т-хелперов в сторону Th2-клеток.
Учитывая, что во фракции лимфоцитов находятся Т-, В-лимфоциты, NК-клетки, присутствие которых может оказывать влияние на конечный результат, дальнейшие эксперименты проводились с использованием CD4+-клеток, основных продуцентов IL-4. Как видно из рис. 13, b-эндорфин усиливает продукцию IL-4 во фракции мононуклеаров и не влиет на уровень IL-4в культуре CD4+-клеток. Добавление к CD4+-лимфоцитам моноцитов приводит к восстановлению уровня продукции IL-4 под воздействием b-эндорфина. Аналогичный по силе и направленности эффект на продукцию IL-4 CD4+-лимфоцитами оказывает селективный d-агонист DADLE (рис. 14). Анализ влияния m-агониста DAGO выявил тенденцию
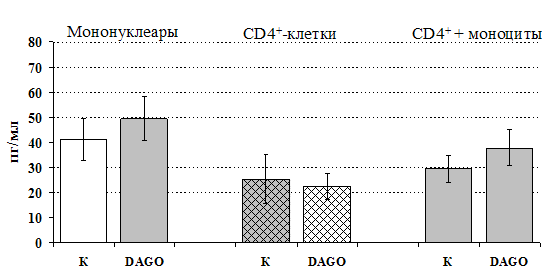
Рис 15. Влияние DAGO 10-8 М на продукцию IL-4 фракцией мононуклеаров, CD4+-лимфоцитами и CD4+-лимфоцитами в присутствии моноцитов в присутствии ФГА 2,5 мкг/мл.
* - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Стъюдента.
к усилению продукции IL-4 фракцией мононуклеаров (рис. 15), однако статистически достоверного эффекта достичь не удалось. Как видно из рис. 16, внесение b-эндорфина на ФГА-индуцированную продукцию IL-2 мононуклеарами, CD4+-лимфоцитами и комбинацией CD4+-лимфоциты+моноциты влияния не оказывает. Таким образом, полученные данные свидетельствуют о том, что от присутствия моноцитов зависит направленность влияния b-эндорфина и d-агониста DADLE на Th1/Th2-поляризацию лимфоцитов. Учитывая важное участие d-рецепторов в регуляции синтеза IL-4 была проанализирована их роль в эффекте b-эндорфина на продукцию данного цитокина. В условиях блокады d-рецепторов нивелируется усиливающее действие пептида на уровень IL-4 в нефракционированных клеточных культурах. Во фракции лимфоцитов b-эндорфин и налтриндол на продукцию IL-4 не влияют (рис. 17).
Таким образом, результаты проведённых исследований свидетельствуют о важной роли моноцитов в регуляции секреторной активности клеток адаптивного иммунитета, при этом как агонисты, так и антагонисты опиатных рецепторов оказывают самостоятельные эффекты на активность клеточных популяций.
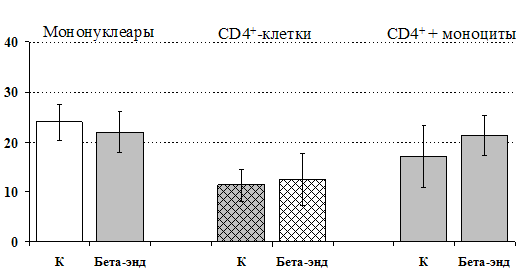
Рис 16. Влияние b-эндорфина 10-7 М на продукцию IL-2 фракцией мононуклеаров, CD4+-лимфоцитами и CD4+-лимфоцитами в присутствии моноцитов в присутствии ФГА 2,5 мкг/мл.
* - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Стъюдента.
А Б
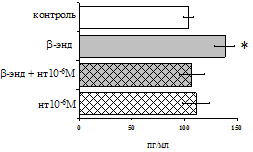
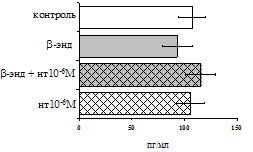
Рис. 17. Эффекты b-эндорфина на фоне блокады d-опиатных рецепторов на продукцию IL-4 в нефракционированной клеточной взвеси (А) и фракции лимфоцитов (Б) в присутствии ФГА 2,5 мкг/мл.
* - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Стъюдента.
Роль b-эндорфина в регуляции фагоцитарной активности клеток естественного звена иммунитета. По нашим данным, β-эндорфин в концентрациях 10-7 и 10-8 М (46,25±1,57 в контроле – 51,3±1,66 - β-эндорфин 10-7 М; P<0,001 к контролю) увеличивает процент фагоцитоза и фагоцитарное число нейтрофилов (0,61±0,02 в контроле - 0,68±0,03 β-эндорфин 10-7 М; P<0,001 к контролю). Помимо нейтрофилов пептид усиливает фагоцитарную активность моноцитов в концентрациях 10-7 – 10-8 М, увеличивая процент фагоцитоза (36,9±2,75 в контроле – 47,7±4,8 - β-эндорфин 10-7 М; P<0,05 к контролю) и фагоцитарное число (0,44±0,03 в контроле - 0,67±0,08 β-эндорфин 10-7 М; P<0,05 к контролю). Так же b-эндорфин стимулирует общий (суммарный) фагоцитоз. На фагоцитарную активность эозинофилов β-эндорфин не влияет.
Роль b-эндорфина в регуляции цитокинпродуцирующей функции моноцитов и нейтрофилов. Как видно из табл. 5, 6, ЛПС усиливает продукцию IL-1b, TNF-a, IL-6 только в культурах с фракцией моноцитов, в то время как в культуре лейкоцитов эффект ЛПС на синтез IL-1b, TNF-a, IL-6 отсутствует. В тоже время уровень IL-8 в ответ на ЛПС усиливается в нефракционированной клеточной фракции и не изменяется в очищенной моноцитарной фракции. В нефракционированной клеточной культуре b-эндорфин (10-7-10-11М) активирует LPS-индуцированную продукцию IL-1b, не влияя на синтез IL-6, TNF-a и угнетая продукцию IL-8 в концентрациях 10-7 и 10-11М. b-эндорфин в концентрациях 10-7 - 10-11М усиливает продукцию IL-1ra, рецепторного антагониста IL-1b. Значительно менее выраженный стимулирующий эффект пептид оказывает на спонтанную продукцию IL-1b в концентрациях 10-7 и 10-9М. На индуцированную субоптимальной дозой ФГА продукцию исследуемых цитокинов, а так же на их спонтанный и ЛПС-индуцированный синтез в очищенной фракции моноцитов b-эндорфин не влияет.
Данные, представленные на рис. 18 указывают на отсутствие отмены стимулирующего эффекта b-эндорфина на уровень IL-1b в условиях блокады опиатных рецепторов неселективным антагонистом налоксоном (d, m) и селективным d-антагонистом налтриндолом в течение 24 ч культивирования. Также обнаруживается самостоятельный стимулирующий эффект налтриндола на продукцию IL-1b. Выявленная динамика сохраняется в течение 48 ч культивирования. Действие b-эндорфина на продукцию антагониста IL-1b IL-1ra (рис. 19) имело картину, схожую с полученной нами при анализе продукции IL-1b. b-эндорфин и налтриндол
Таблица 5. Влияние b-эндорфина на продукцию IL-1b, TNF-a, IL-6 в нефракционированной лейкоцитарной суспензии
| Цитокин, |
Экспериментальное |
Концентрация b-эндорфина, M |
|||
| пг/мл | воздействие | контроль |
10-7 |
10-9 |
10-11 |
| IL-1b, |
Без индуктора |
193,01± | 271,66± | 266,57± | 238,22± |
| n=8 | 39,16 | 77,96* | 49,18* | 50,19 | |
| ЛПС 0,1 мкг/мл | 190,87± | 305,76± | 300,95± | 279,41± | |
| 54,43 | 49,50*** | 76,95** | 62,40* | ||
| TNF-a, | Без индуктора | 253,72± | 286,79± | 277,46± | 290,90± |
| n=8 | 52,60 | 61,43 | 67,08 | 62,32 | |
| ЛПС 0,1 мкг/мл | 269,59± | 297,93± | 295,42± | 295,49± | |
| 53,90 | 66,45 | 68,59 | 60,45 | ||
| IL-6, | Без индуктора | 1115,41± | 1084,09± | 1101,83± | 1109,70± |
| n=8 | 30,54 | 50,23 | 56,55 | 46,66 | |
| ЛПС 0,1 мкг/мл | 1094,73± | 1082,85± | 1051,07± | 1115,41± | |
| 36,40 | 28,72 | 36,16 | 33,88 | ||
| IL-8, | Без индуктора | 1353,68± | 1340,28± | 1664,63± | 1313,30± |
| n=4 | 114,42 | 104,42 | 44,10 | 156,25 | |
| ЛПС 0,1 мкг/мл | 1699,20± | 1278,40± | 1635,43± | 1364,28± | |
| 68,25а | 31,34*** | 68,31 | 59,85** | ||
| IL-1ra, | Без индуктора | 1811,38± | 1840± | 1817,25± | 1932,5± |
| n=4 | 272,30 | 275,54 | 431,75 | 297,55 | |
| ЛПС 0,1 мкг/мл | 1957,13± | 2346,5± | 2168,13± | 2175,13± | |
| 232,16 | 198,97*** | 261,74* | 198,06* | ||
Примечание. Здесь и в табл. 6: * - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Фишера наименьшей значимой разницы, a – р <0,05 – к спонтанной продукции.
стимулируют продукцию IL-1ra по сравнению с контролем как на 1-е, так и на 2-е сутки культивирования. Отмены стимулирующего влияния пептида на фоне блокады опиатных рецепторов не наблюдается. Анализ влияния пептида на продукцию IL-8 показал статистически достоверный угнетающий эффект как на 24, так и на 48 ч культивирования. В условиях блокады опиатных рецепторов налоксоном и налтриндолом угнетающее влияние b-эндорфина нивелируется. Внесение налоксона и налтриндола на продукцию IL-8 не влияет.
Таблица 6. Влияние b-эндорфина на продукцию IL-1b, TNF-a, IL-6 в очищенной фракции моноцитов
| Цитокин, |
Экспериментальное |
Концентрация b-эндорфина, M |
|||
| пг/мл | воздействие | контроль |
10-7 |
10-9 |
10-11 |
Без индуктора |
153,22± | 121,65± | 249,92± | 118,71± | |
| IL-1b, | 33,48 | 19,83 | 84,88 | 28,97 | |
| n=8 | ЛПС 0,1 мкг/мл | 230,26± | 202,84± | 223,97± | 235,21± |
| 42,25а | 36,47 | 71,16 | 53,29 | ||
| Без индуктора | 137,52± | 153,81± | 148,31± | 133,20± | |
| TNF-a, | 51,33 | 58,24 | 57,56 | 55,31 | |
| n=8 | ЛПС 0,1 мкг/мл | 163,21± | 178,81± | 174,97± | 178,73± |
| 60,48а | 69,78 | 68,63 | 70,21 | ||
| Без индуктора | 1204,61± | 1174,58± | 1281,19± | 1243,46± | |
| IL-6, | 354,15 | 356,34 | 357,40 | 332,03 | |
| n=8 | ЛПС 0,1 мкг/мл | 1435,55± | 1442,89± | 1346,26± | 1414,19± |
| 379,02а | 340,04 | 342,73 | 367,26 | ||
| Без индуктора | 1198,38± | 1230,63± | 1294,63± | 1148,68± | |
| IL-8, | 153,25 | 85,79 | 219,75 | 200,81 | |
| n=4 | ЛПС 0,1 мкг/мл | 1307,22± | 1422,48± | 1159,64± | 1228,32± |
| 213,04 | 90,89 | 121,80 | 154,70 | ||
| Без индуктора | 280,38± | 364,25± | 352,83± | 359,05± | |
| IL-1ra, | 90,82 | 128,06 | 133,52 | 146,50 | |
| n=4 | ЛПС 0,1 мкг/мл | 302,13± | 343,40± | 377,35± | 348,80± |
| 88,60 | 115,95 | 148,54 | 141,58 | ||
При оценке влияния опиатов на уровень TNF-a выявлено угнетение продукции TNF-a по сравнению с контролем при совместном внесение в культуры b-эндорфина и антагонистов опиатных рецепторов на 24 и 48 ч культивирования (см. рис. 18). На продукцию IL-6 на 24 ч культивирования b-эндорфин не влияет. На 48 ч культивирования продукцию IL-6 стимулирует налтриндол как при изолированном внесении, так и в комбинации с b-эндорфином. Учитывая, что во фракции лейкоцитов помимо моноцитов, основных продуцентов провоспалительных цитокинов, присутствуют гранулоциты, преимущественно нейтрофилы (Abraham et al., 2003; Fujiharaa et al., 2003; Xing, Remick, 2004), так же способные продуцировать IL-1b и IL-8, мы
24 ч 48 ч
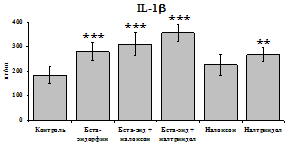
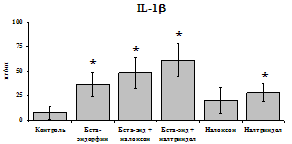
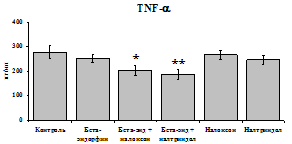
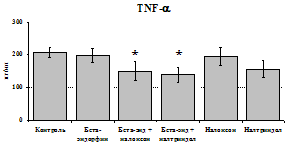

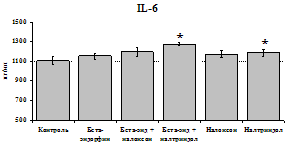
Рис. 18. Влияние b-эндорфина 10-7М в условиях блокады опиатных рецепторов на продукцию IL-1b, TNF-a, IL-6 в нефракционированной клеточной суспензии в присутствии ЛПС.
* - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Стъюдента..
исследовали влияние b-эндорфина на продукцию IL-1b и IL-8 во фракциях мононуклеаров и нейтрофилов.
Результаты исследований влияния b-эндорфина на продукцию IL-1 и IL-8 во фракциях мононуклеаров и нейтрофилов представлены на рис. 20. b-эндорфин 10-7М стимулирует спонтанную и ЛПС-индуцированную продукцию IL-1b мононуклеарами. Аналогичная картина наблюдается во фракции нейтрофилов. На фоне LPS наблюдается статистически значимое усиление продукции IL-1b. b-эндорфин 10-7М стимулирует как спонтанную, так и LPS-индуцированную продукцию IL-1b нейтрофилами. ЛПС-индуцированная продукция IL-8 мононуклеарами и нейтрофилами под воздействием b-эндорфина снижается. В культурах без ЛПС пептид уровень IL-8 не изменяет.
При анализе влияния b-эндорфина на продукцию исследуемых цитокинов мононуклеарами в присутствии ФГА были получены следующие результаты (рис. 21). Во фракции мононуклеаров ФГА, как и ЛПС, усиливает выработку IL-1b и не влияет на уровень IL-8. b-эндорфин стимулирует как спонтанную, так и ФГА-индуцированную продукцию IL-1b мононуклеарами. Уровень IL-8 под воздействием пептида в нестимулированных ФГА пробах не изменяется, а в стимулированных - снижается. Как следует из полученных данных, продукцию цитокинов семейства IL-1 нефракционированными лейкоцитами, нейтрофилами и мононуклеарами (но не очищенными моноцитами) b-эндорфин активирует, а продукцию IL-8 угнетает, таким образом, оказывая двоякое действие, с одной стороны, активируя процессы пролиферации и дифференцировки, а, с другой, угнетая хемотаксис. При этом в отличие от IL-1, депрессия IL-8
24 ч 48 ч
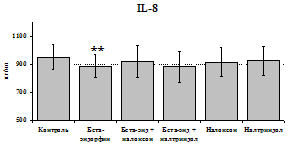
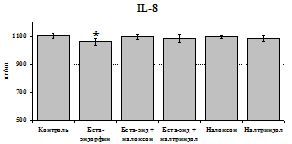
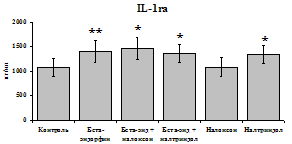
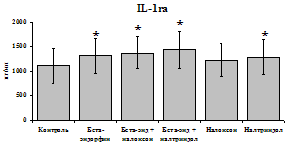
Рис. 19. Влияние b-эндорфина 10-7М в условиях блокады опиатных рецепторов на продукцию IL-8 и IL-1ra в нефракционированной клеточной суспензии в присутствии ЛПС.
* - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Стъюдента..
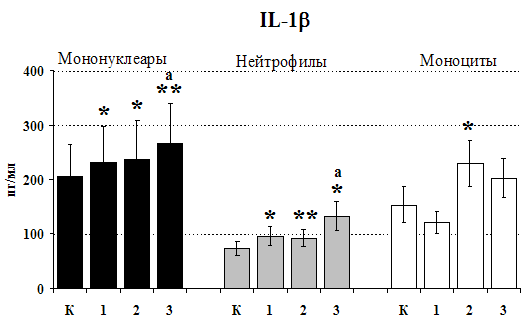
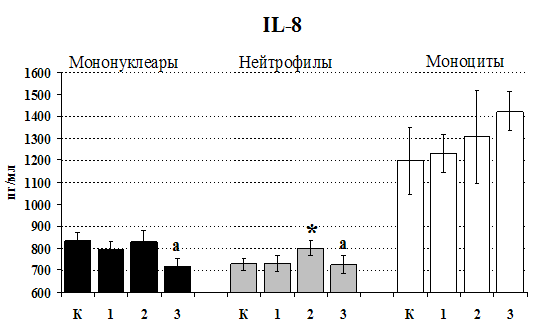
Рис. 20. Влияние b-эндорфина 10-7М на продукцию IL-1b и IL-8 во фракциях мононуклеаров, нейтрофилов и моноцитов, стимулированных ЛПС.
К контроль, 1 - b-эндорфин, 2 – ЛПС, 3 – ЛПС+b-эндорфин. * - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю, а - р < 0,05 к ЛПС по парному t-критерию Стъюдента..
проявляется только в ЛПС и ФГА-индуцированных культурах и нивелируется опиатными антагонистами, что указывает на вовлечённость различных рецепторных механизмов в регуляцию синтеза IL-1b и IL-8 b-
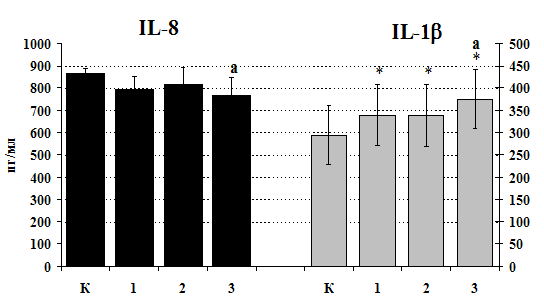
Рис. 21. Влияние b-эндорфина 10-7М на продукцию IL-1b и IL-8 мононуклеарами, стимулированными ФГА 2,5 мкг/мл в течении 24 ч культивирования.
К контроль, 1 - b-эндорфин, 2 – ФГА, 3 – ФГА+b-эндорфин. * - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю, а - р < 0,05 к ФГА по парному t-критерию Стъюдента.
эндорфином. Ранее (Nandhra, 2000) был зарегистрирован угнетающий блокируемый налоксоном, эффект b-эндорфина на продукцию IL-8 клетками хориодецидуальной оболочки. Н.Л. Елизарова с соавт. (2001) установили, что аминокислотная последовательность β-эндорфина в молекуле препарата тимуса – тактивина обусловливает активацию функциональной активности лимфоцитов, усиление фагоцитоза и снижение способности моноцитов к адгезии, последняя функция отменялась налоксоном, в то время как усиление фагоцитоза налоксоном не отменялось. По нашим данным, депрессия продукции IL-8 отменяется селективным d-антагонистом налтриндолом, что свидетельствует о возможности реализации эффектов b-эндорфина через d-рецептор. В то же время, как показано в работе B.M. Sharp (2006), в зависимости от экспериментальной модели и объекта исследования d-рецептор может опосредовать проведение как стимулирующих, так и угнетающих сигналов.
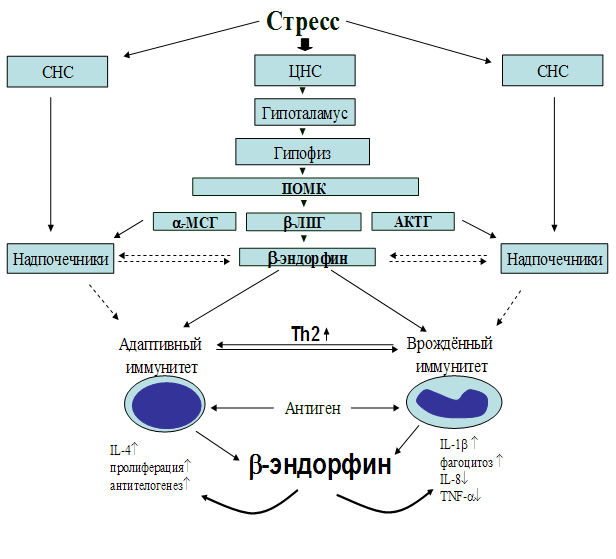
Рис. 22. Схема возможного механизма влияния b-эндорфина на процессы иммуногенеза.
Заключение
Результаты проведённых исследований представлены в виде схемы возможного механизма влияния b-эндорфина на процессы иммуногенеза (рис. 22). Установлено, что b-эндорфин является важным фактором в поддержании внутреннего гомеостаза. Приведённые в данные, свидетельствуют о том, что b-эндорфин поддерживает гомеостаз иммунной системы через балансирование Th1 и Th2 ответа, смещая поляризацию Т-хелперов в сторону Th2-клеток, параллельно усиливая естественную резистентность. Следовательно, b-эндорфин является кофактором, опосредующим переключение дифференцировки Т-хелперов с Th1 на Th2 тип и играющим решающую роль при целом ряде иммунопатологических состояний, а иммуномодулирующие функции опиоидных пептидов могут представлять значительный терапевтический интерес.
Оценивая роль эндогенных нейропептидов в стрессиндуцированных изменениях иммунной системы необходимо отметить, что полученные данные прямо указывают на непосредственное участие эндогенных опиатов в активации иммунной системы. Направленность ряда физиологических эффектов эндогенных опиоидных пептидов при стрессе противоположна эффектам глюкокортикоидов и катехоламинов. Ранее глюкокортикоиды и катехоламины рассматривались исключительно как стрессреализующие факторы, в то же время их основная биологическая роль может заключаться в ограничении опасной для организма стрессиндуцированной активации иммунных процессов.
ВЫВОДЫ
1. Подтверждена интегральная роль эндогенной опиоидной системы в нейроэндокринной регуляции иммуногенеза в норме, введении глюкокортикоидов и катехоламинов и в условиях стрессорного воздействия. Основные компоненты эндогенной опиоидной системы принимают непосредственное участие в активации иммунной системы при стрессе.
2. Установлено, что в условиях блокады m, d-опиатных рецепторов в период индукции иммунного ответа повышается степень выраженности иммуносупрессивного действия гормонов стресса (глюкокортикоидов и катехоламинов) на антителогенез и регистрируется снижение степени выраженности угнетающего действия глюкокортикоидов на количество ядросодержащих клеток в регионарном лимфатическом узле.
3. Впервые показано, что b-эндорфин в высоких (100 мкг/мл) дозах угнетает, в низких (0,0005 мкг/кг) дозах стимулирует гуморальное звено иммунитета и при этом не влияет на степень выраженности реакции гиперчувствительности замедленного типа. В клеточных культурах b-эндорфин стимулирует пролиферацию лимфоцитов и продукцию IL-4, не влияет на синтез IL-2, IFN-g и при этом усиливает степень поляризации Т-хелперов в направлении Th2-клеток. Доминирующая роль в реализации стимулирующих эффектов b-эндорфина на пролиферацию и продукцию IL-4 принадлежит d-рецепторам.
4. Впервые обнаружена зависимость стимулирующего эффекта опиоидных пептидов на пролиферацию лимфоцитов и продукцию IL-4 от присутствия моноцитов в клеточной культуре.
5. Показано, что b-эндорфин стимулирует фагоцитарную активность клеток естественного иммунитета и оказывает модулирующее действие на цитокинпродуцирующую функцию моноцитов и нейтрофилов. В условиях блокады m, d-опиатных рецепторов отмены стимулирующего влияния b-эндорфина на продукцию IL-1b не наблюдается, при этом угнетение продукции IL-8 опиатными антагонистами отменяется.
Список работ, опубликованных по теме диссертации
Экспериментальные статьи
1. Шилов Ю.И., Гейн С.В. Адренергическая регуляция пролиферативного ответа лимфоцитов в культурах с Т-клеточными митогенами // Бюллетень экспериментальной биологии и медицины. - 1999. - Т. 128, № 8. - С. 207-209.
2. Шилов Ю.И., Гейн С.В, Черешнев В.А. Влияние блокады b-адренергических рецепторов при стрессе на антителообразование, гиперчувствительность замедленного типа, функции фагоцитирующих клеток при локальной форме иммунного ответа // Russian Journal of Immunology. - 2001. - V. 6, № 3. - P. 301-308.
3. Гейн С.В., Симоненко Т.А., Черешнев В.А. Влияние b-эндорфина и селективного агониста m-опиатных рецепторов DAGO на пролиферативную активность лимфоцитов // Доклады академии наук. - 2003. - Т. 391, № 1. - С. - 127-129.
4. Гейн С.В., Симоненко Т.А., Тендрякова С.П. Влияние агонистов опиатных рецепторов на процесс антителообразования и гиперчувствительность замедленного типа при локальном иммунном ответе // Вестн. Пермск. университета. Серия Биолгия. - 2004. - Вып. 2. - С. 166-168.
5. Гейн С.В., Тендрякова Т.А. Иммуномодулирующие эффекты гидрокортизона и адреналина в условиях блокады опиатных рецепторов // Докл. Академии наук. - 2004. - Т. 399, № 3. - С. 412-414.
6. Gein S.V., Simonenko T. A., Tendryakova S. P. The effects of rotation stress on measures of immunity. The role of opiate receptors // Neuroscience and Behavioral Physiology. - 2004. - V. 34, № 9. - P. 935-938.
7. Гейн С.В., Баева Т.А. Роль опиоидных пептидов в регуляции пролиферации лимфоцитов и изменении Тh1/Тh2 цитокинового профиля // Проблемы эндокринологии. - 2005. - № 5. - С. 49-51.
8. Гейн С.В., Шаравьёва И.Л, Баева Т.А., Тендрякова Т.А. Роль блокады различных типов опиатных рецепторов в регуляции иммуномодулирующих эффектов ротационного стресса // Вестник Уральской медицинской академической науки. - 2005. - № 3. - С. 70-73.
9. Гейн С.В., Баева Т.А., Гейн О.Н., Черешнев В.А. Роль моноцитов в реализации эффектов b-эндорфина и селективных агонистов m- и d-опиатных рецепторов на пролиферативную активность лимфоцитов периферической крови // Физиология человека. - 2006. - Т. 32, № 3. С. 111-116.
10. Гейн С.В., Шаравьёва И.Л, Тендрякова Т.А. Роль d-опиатных рецепторов в регуляции гуморального и клеточноопосредованного иммунного ответа при ротационном стрессе // Доклады Академии наук. - 2006. - Т. 407, № 1. - С. 127-129.
11. Гейн С.В., Баева Т.А., Кичанова О.А. Влияние b-эндорфина на антителогенез и продукцию ИЛ-4 в условиях блокады опиатных рецепторов // Бюллетень экспериментальной биологии и медицины. - 2006. - Т. 142, № 8. – С. 192-195.
12. С.В. Гейн, Е.Г. Чижова, С.П. Тендрякова Влияние гидрокортизона и адреналина в условиях блокады m- и d-опиатных рецепторов на развитие локального иммунного ответа у мышей // Российский физиологический журнал им. Сеченова. - 2006. - Т. 92, № 7. - С. 827-834.
13. Горшкова К.Г., Гейн С.В., Тендрякова С.П. Роль бета-эндорфина в регуляции продукции IL-1b и IL-8 моноцитами и нейтрофилами периферической крови // Вестник Уральской медицинской академической науки. - 2006. - № 3-1(14). - С. 46-47.
14. Гейн С.В., Баева Т.А., Чижова Е.Г., Кичанова О.А., Шаравьева И.Л. Эндогенная опиоидная система и гормоны стресса в сочетанной регуляции иммунных реакции при остром стрессе // Вестник Уральской медицинской академической науки. - 2006. - № 3-1(14). - С. 38-40.
15. Гейн С.В., Горшкова К.Г., Тендрякова С.П.. Роль b-эндорфина в регуляции продукции провоспалительных цитокинов моноцитами периферической крови in vitro // Бюллетень экспериментальной биологии и медицины. - 2007. - Т. 143, № 2. - С. 175-178.
Труды конференций
16. Shilov Ju.I., Gein S.V., Chereshnev V.A.. Adrenergic and cAMP-dependent regulation of mitogen-induced lymphocyte proliferative responses // Annual Meeting of AAAAI. J. Allergy and Clin. Immunol. - 1998. - V. 101, № 1. - S. 38-38.
17. Gein S.V., Shilov Ju.I. Influence of stress on expression of immune reactions // Межд. конф. "Проблемы загрязнения окружающей среды-98". Москва, 1998. - С. 238-238.
18. Шилов Ю.И., Гейн С.В. Влияние блокады бета-адренергических рецепторов и иммобилизационного стресса на реакции клеточноопосредованного и гуморального иммунитета // Научная сессия Пермской государственной медицинской академии. Пермь, 1998. - С. 68-68.
19. Гейн С.В., Шилов Ю.И. Адренергическая и цАМФ-зависимая регуляция пролиферативного ответа лимфоцитов в культурах с митогенами // Научная сессия Пермской государственной медицинской академии. Пермь, 1998. - С. 69-69.
20. Гейн С.В., Шилов Ю.И. Адренергическая и цАМФ-зависимая модуляция пролиферативного ответа лимфоцитов в культурах с митогенами // V Российский национальный конгресс "Человек и лекарство". Москва, 1998. - C. 330-330.
21. Шилов Ю.И., Гейн С.В. Модуляция пропранололом стресс-индуцированных изменений иммунных реакций // Там же. - C. 340-340.
22. Гейн С.В., Шилов Ю.И., Черешнев В.А. Влияние адренергических соединений на пролиферативный ответ лимфоцитов // II-й Национальный конгресс Российской Ассоциации аллергологов и клинических иммунологов «Современные проблемы аллергологии, клинической иммунологии и иммунофармакологии». Москва, 1998. - С. 345-345.
23. Шилов Ю.И., Гейн С.В. Влияние иммобилизационного стресса и блокады b-адренергических рецепторов на антителообразование и реакции гиперчувствительности замедленного типа в регионарных лимфатических узлах и селезенке при подкожной иммунизации // Иммунология. - 1998. - № 6. - С. 25-26.
24. Шилов Ю.И., Гейн С.В. Влияние адреналина и селективных антагонистов a1- и a2-адренорецепторов на пролиферативный ответ лимфоцитов периферической крови // Матер. III Всерос. научного Форума «Дни иммунологии в Санкт-Петербурге». Медицинская иммунология. - 1999. - Т. 1, № 3-4. - С. 28-29.
25. Гейн С.В., Шилов Ю.И. Роль адренорецепторов в регуляции пролиферативного ответа лимфоцитов в культурах с Т-клеточными митогенами // Всероссийской научно-практической конференция "Актуальные вопросы эндокринологии". Пермь, 1999. - С. 101-102.
26. Шилов Ю.И., Гейн С.В., Орлова Е.Г., Черешнев В.А. Влияние блокады b-адренергических рецепторов на иммунный ответ и функциональную активность фагоцитирующих клеток в условиях стресса // Аллергология и иммунология. - 2000. - Т. 1, № 2. - С. 116-117.
27. Шилов Ю.И., Гейн С.В. Иммуномодулирующие эффекты адренергических соединений в системах in vivo и in vitro // Труды российской научной конференции «Факторы клеточного и гуморального иммунитета при различных физиологических и патологических состояниях». Челябинск, 2000. - С. 134-135.
28. Кеворков Н.Н., Шилов Ю.И., Бахметьев Б.А., Ширшёв С.В., Куклина Е.М., Гейн С.В., Сидоров Д.В., Заморина С.А., Орлова Е.Г., Лихачева Н.С., Груздева Е.А., Харитонова А.В. Нейроэндокринная регуляция некоторых функций фагоцитирующих и иммунокомпетентных клеток // Тез. докл. научной конференции «Актуальные проблемы фундаментальных исследований в области биологии и медицины». Санкт-Петербург, 2000. - С. 67-68.
39. Гейн С.В. Роль b-адренергических механизмов в регуляции реакций клеточноопосредованного и гуморального иммунного ответа при стрессе // II конференция молодых ученых России с международным участием “Фундаментальные науки и прогресс клинической медицины”. Москва. - Т. 1. - С. 191-191.
30. Гейн С.В., Симоненко Т.А., Тендрякова С.П., Шилов Ю.И. Влияние блокады d-,m-опиатных рецепторов при стрессе на показатели клеточноопосредованного, гуморального иммунитета в условиях развития локальной формы иммуного ответа // I конференция иммунологов Урала. Екатеринбург, Иммунология урала. – 2001. - №1. – С. 2-3.
31. Гейн С.В., Симоненко Т.А., Тендрякова С.П. Участие d-, m, k-опиатных рецепторов в развитии реакции гиперчувствительности замедленного типа и изменении числа антителообразующих клеток в модели принудительного головокружения // Труды международной научной конференции «Перспективы развития естественных наук в высшей школе». Пермь, 2001. - Т. 3. - С.120-124.
32. Гейн С.В., Симоненко Т.А., Тендрякова С.П. Роль d-,m-,k-опиатных рецепторов в регуляции функций иммунокомпетентных и фагоцитирующих клеток при стрессе // II конференция иммунологов Урала. Пермь, Иммунология Урала. – 2002. – №1. – С. 4.
33. Гейн С.В., Симоненко Т.А. Участие b-эндорфина и селективных лигандов d, m-опиатных рецепторов в регуляции пролифератив-ной активности лимфоцитов in vitro // III конференция иммунологов Урала. Челябинск, Иммунология урала. – 2003. - № 1. - С. 5.
34. Гейн С.В., Симоненко Т.А. Роль b-эндорфина и опиодных пептидов в регуляции пролиферации лимфоцитов и изменении Тh1/Тh2 цитокинового профиля // Матер. VIII Всерос. научного Форума с международным участием «Дни иммунологии в Санкт-Петербурге». Медицинская иммунология. - 2004. - V. 6, № 3-5. - P. 226-227.
35. Гейн С.В., Симоненко Т.А. Опиоидная регуляция пролиферативного ответа лимфоцитов in vitro // Rus. J. Immunol. - 2004. - V. 9, № 1. - P. 38-38.
36. Гейн С.В., О.Н. Гейн, Т.В. Гаврилова Механизмы стрессорных нарушений функций иммунной системы и их коррекция // XIX Съезд физиологического общества им. И.П. Павлова 19-24 сентября 2004. Тез. докл. – Российский физиологический журнал им. Сеченова. - 2004. - Т. 90, №8. - С. 106-107.
37. Гейн С.В., Чижова Е.Г. Влияние гидрокортизона на фоне блокады опиатных рецепторов на развитие локального иммунного ответа // IX международная Пущинская школа-конференция молодых ученых «Биология-наука XXI века». Пущино, 2005. - С. 174.
38. Гейн С.В., Баева Т.А. Роль опиодных пептидов в регуляции пролиферации лимфоцитов и изменении Th1/Th2 цитокинового профиля // Там же. - С. 69.
39. Баева Т.А., Гейн С.В., Шаравьева И.Л., Тендрякова С.П. Роль блокады различных типов опиатных рецепторов в регуляции иммуномодулирующих эффектов ротационного стресса // Международная конференция "Проблемы загрязнения окружающей среды-2005". Пермь, 2005. - С. 56.
40. Гейн С.В., Баева Т.А., Черешнев В.А. Лиганды опиатных рецепторов в регуляции пролиферативной активности лимфоцитов in vitro // Международная конференция "Проблемы загрязнения окружающей среды-2005". Пермь, 2005. - С. 94.
41. Шилов Ю.И., Шилов С.Ю., Ланин Д.В., Гейн С.В., Орлова Е.Г., Черешнев В.А. Адренергические механизмы регуляции иммунного ответа и функций неспецифических эффекторных клеток при стрессе // Матер. IX Всерос. научного Форума с международным участием «Дни иммунологии в Санкт-Петербурге». Медицинская иммунология. - 2005. - Т. 7, № 2-3. - С. 127-127.
42. Гейн С.В., Баева Т.А., Черешнев В.А. Роль моноцитов в опиатэргической регуляции процессов пролиферации лимфоцитов и цитокинового синтеза // Там же. - С. 110-110.
43. Гейн С.В., Баева Т.А., Черешнев В.А. Роль b-эндорфина в регуляции процессов пролиферации и продукции IL-4 лимфоцитами периферической крови in vitro // IV конференция иммунологов Урала. Уфа, Иммунология Урала. - 2005. - № 1 (4). - С. 4-5.
44. Гейн С.В., Чижова Е.Г., Тендрякова С.П. Влияние гидрокортизона на фоне блокады d-, m-опиатных рецепторов на иммунный ответ // Там же. - С. 5-6.
45. Гейн С.В., Горшкова К.Г. Влияние b-эндорфина на продукцию IL-1b, TNF-a, IL-6 моноцитами периферической крови // V конференция иммунологов Урала. Оренбург, Иммунология урала. - 2005. - № 1. - с. 6-7.
46. Gein S.V., Gorshkova K.G., Baeva T.A. b-endorphin-induced modulation of cytokine production by peripheral blood leukocytes // International symposium «Interaction of the nervous and immune systems in health and disease» Saint-Petersburg, 2007. - Р. 26-27.
Список основных сокращений
АКТГ - адренокортикотропный гормон
АОК - антителообразующая клетка
ГЗТ - гиперчувствительность замедленного типа
ЛПС - липополисахарид
ПОМК - проопиомеланокортин
РБТЛ - реакция бласттрансформации лимфоцитов
СНС - симпатическая нервная система
ФГА - фитогемагглютинин
ЦНС - центральная нервная система
ЯСК - ядросодержащая клетка
CD - маркеры дифференцировки лимфоцитов
DADLE - δ-агонист ([d–Ala2,d-Leu5] - энкефалин
DAGO - m-агонист ([d-Ala2,N-Me-Phe4,Gly5-ol]-энкефалин
IFN - интерферон
IL - интерлейкин
MPF - меланотропин потенцирующий фактор
TNF - фактор некроза опухоли
© 2009 База Рефератов