
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Учебное пособие: Нервова тканина
Учебное пособие: Нервова тканина
Модуль 3. Міогістологія, нейрогістологія
Лекція № 8. Тема: Нервова тканина
Зміст
1. Гістогенез нервово тканини
2. Нервові клітини
3. Нейроглія
4. Нервов волокна
5. Синапси
6. Нервов закінчення
Нервова тканина - це високодиференційована спеціалізована тканина, яка формує інтегруючу систему організму — нервову. Її структури здатні сприймати подразнення, трансформувати його в нервовий імпульс, швидко його передавати, зберігати інформацію, синтезувати біологічно активні речовини. Завдяки цьому нервова система регулю взаємозв'язок органів і систем організму та адаптацію його до екологічних умов середовища. Нервова тканина складається із двоякого роду органічно зв'язаних клітинних елементів: нервових клітин (нейронів, нейроцитів), здатних утворювати нервове збудження та проводити нервовий імпульс; нейроглії — клітини забезпечують опорну, трофічну, розмежувальну, секреторну та захисну функції.
1. Гістогенез нервово тканини
Нервова тканина розвивається із нервової пластинки, яка є потовщенням ектодерми. Нервова пластинка послідовно диференціюється у нервовий жолобок і нервову трубку, яка, замикаючись, відокремлюється від шкірної ектодерми. В наступні періоди ембріогенезу з нервової трубки утворюється головний та спинний мозок. Частина клітин нервової пластинки залишається поза нервовою трубкою і утворює нервовий гребінь або гангліозну пластинку. Клітини гребеня мігрують у латеральному та вентральному напрямках і дають такі похідні, як ядра черепних нервів, нейрони спинальних та симпатичних гангліїв, лемоцити, меланоцити шкіри, клітини АРИД-системи. Клітини нервової трубки — нейроепітеліальні або вентрикулярні розміщуються у кілька рядів, їх апікальний полюс спрямований у порожнину нервової трубки, а базальний — контактує із субпіальною мембраною. Здатність цих клітин до розмноження зменшується у процесі ембріогенезу і після народження втрачається зовсім.
Морфологічно подібн нейроепітеліальні, вентрикулярні клітини шляхом диференціації перетворюються у різні типи клітин нервової тканини. Частина із них дає початок нейронам, інша гліальним клітинам (епендимоцитам, астроцитам, олігодендроцитам).
2. Нервові клітини
Нервові клітини (нейроцити, нейрони) є морфологічними та функціональними одиницями нервово тканини. Нейроцити різних відділів нервової системи є специфічними за розмірами формою. Складаються нейрони із тіла (перикаріону) і відростків. Наявність останніх - найхарактерніша ознака нервових клітин. Самі відростки забезпечують проведення нервового мпульсу на різну відстань від мікрометрів до метра і більше, а також забезпечують зв'язок нейронів у складі рефлекторних дуг. Нейрони, не здатні до мітотичного поділу, мають тривалий життєвий цикл, термін їх існування збігається із терміном життя індивідума. Розміри перикаріону нейронів дуже різноманітні — від 4—6 мкм (клітини — зерна мозочка) до 120—130 мкм (гігантські пірамідні клітини півкуль головного мозку). Відростки нейроцитів за функціональним значенням поділяють на аксони і дендрити.
Аксон — (нейрит) це довгий відросток, який завжди в клітині один. Діаметр по всій довжин незмінний, він не розгалужується, але може давати колатералі, що мають інший напрямок. Закінчується аксон термінальним розгалуженням, по ньому проходить нервовий імпульс у напрямку від перикаріону нейрона.
Дендрити — це, здебільш, короткі відростки, які розгалужуються деревоподібно («дендрон» — дерево); основи дендритів мають конічне розширення. Нервовий імпульс ці відростки передають у напрямку до тіла нейрона. Нейроцити мають у центрі перікаріону кругле або овальне ядро з незначною кількістю гетерохроматину і значним вмістом еухроматину, що характеризує різний рівень синтетичних процесів, а звідси функціональний стан клітини. Цитоплазма (нейроплазма) нервових клітин характеризується наявністю дуже розвинених органел, що відповідає їх високій функціональній активності. Розрізняють три типи організованих структур нейроплазми: загальні органели, включення та спеціальні органели.
Спеціальними органелами нейронів є хроматофільна субстанція (субстанція Нісля, тигроїд) та нейрофібрили. При світооптичному аналізі хроматофільна субстанція має вигляд зерен різної ведичини, що забарвлюються базофільно (базофільна речовина) і локалізуються у перикаріоні та дендритах. В аксонах та їх початкових сегментах хроматофільну субстанцію ніколи не виявляють. Під електронним мікроскопом цю структуру виявляють гранулярним ендоплазматичним ретикулом з паралельним розміщенням сплющених цистерн (ергастоплазма), де інтенсивно синтезується білок. Хроматофільна субстанція є показником функціонального стану нейрона.
В аксонах, що не містять органел білкового синтезу, цитоплазма постійно переміщується від перікаріона до терміналів. Розрізняють повільне та швидке аксонні транспортування білків ферментів для синтезу медіаторів у синапсах та речовин для синаптичної функції. Ретроградний потік цитоплазми забезпечує повернення ряду компонентів із закінчень у тіло нейрона.
Нейрофібрили можна виявити в нейроплазмі при імпрегнації азотнокислим сріблом. Вони мають вигляд тонких ниток діаметром від 0,4 до 0,6 мкм, які утворюють густу сітку в перикаріоні і мають паралельну орієнтацію у складі дендритів і аксонів, включаючи їх найтонші кінцеві розгалуження. Ультраструктурний аналіз дає змогу віднести нейрофібрили до пучків нейрофіламентів (мікрофіламентів) діаметром від 6 до 10 нм і нейротубул (мікротрубочок діаметром 20—30 нм). Мікрофіламенти мікротрубочки належать до системи цитоскелету нейронів. Структури цитоскелету складаються із білка спектрину, що є аналогом спектрину еритроцитів та тканини головного мозку.
Морфологічна класифікація нейронів грунтується залежно від кількості наявних відростків. За цією ознакою нейрони поділяють на такі різновиди: уніполярні (нейробласти мають єдиний відросток, який є аксоном, що передає нервовий імпульс на другий нейрон); біполярні (мають два відростки — аксон та дендрит), характерні для органів чуття; псевдоуніполярні (мають один відросток, який на певній відстані від перикаріону нейрона поділяється на аксон і дендрит), характерні для спинальних гангліїв; мультиполярні (мають багато відростків, один з яких є аксоном, а вс нші дендритами). Функціональна класифікація нейронів грунтується на положенн нейроцита у складі рефлекторної дуги. Аферентні нейрони (рецепторні, чутливі) сприймають подразнення і трансформують його у нервовий імпульс. Еферентн (моторні, рухові) забезпечують передачу нервового імпульсу на робочу структуру органа. Асоціативні (вставні) нейрони — передають нервовий імпульс між нейронами.
Рефлекторна дуга — це ланцюжок нейронів, який передає нервовий імпульс від чутливого закінчення (рецептора) до рухового (ефектора), розміщеного у робочому органі. Найпростіша рефлекторна дуга складається з двох нейронів: аферентного, дендрит якого закінчується рецептором, а аксон передає імпульс на дендрит еферентного нейрона; еферентного, який своїм аксоном передає імпульс до ефектора у робоч структури органу. Складні рефлекторні дуги містять між аферентними еферентними нейронами кілька асоціативних нейронів. Нервове збудження по рефлекторній дузі передається лише в одному напрямку, названому фізіологічною (або динамічною) полярізацією нейронів. Ізольований нейрон здатний проводити нервовий імпульс в будь-якому напрямку. Односпрямованість передачі імпульсу в межах рефлекторної дуги зумовлена структурною організацією міжнейронного контакту, що названий синапсом.
3. Нейроглія
Нейрони знаходяться у тісному генетичному структурному та функціональному зв'язку з нейроглією. Цей термін був запропонований Р. Вірховим і в буквальному перекладі означа «нервовий клей», а в дійсності це середовище, що оточує нейрони. Складається нейроглія з клітин і виконує опорну, розмежувальну, трофічну, секреторну та захисну функції. Всі клітини нейроглії поділяють на два генетичних види: гліоцити (макроглія) і гліальні макрофаги мікроглія). У свою чергу серед гліоцитів розрізняють епендимоцити, астроцити і олігодендроцити. Макроглія походить, як і нейрони, із матеріалу нервової трубки, а мікроглія — з моноцитів і належить до макрофагічної системи (однак є дані, що мікроглія не має моноцитарного генезу).
Епендимоцити утворюють щільний, епітеліоподібний пласт клітин, які вистеляють спинномозковий канал всі шлуночки мозку. Виникають вони першими у процесі гістогенезу нервово тканини з гліобластів нервової трубки. На цій стадії розвитку епендимоцити виконують розмежувальну й опорну функції. На апікальній поверхні клітини, зверненої у порожнину каналу нервової трубки, утворюється до 40 війок на одну клітину. Після народження війки епендимоцитів поступово зникають і зберігаються лише в деяких ділянках (у водопроводі середнього мозку). Від базального полюсу епендимоцитів відходять довгі відростки, які, розгалужуючись, перетинають усю нервову трубку, утворюючи її опорний апарат. По зовнішній поверхні нервово трубки відростки епендимоцитів утворюють поверхневу гліальну пограничну мембрану, яка відмежовує нервову трубку від інших тканин.
Деякі епендимоцити виконують секреторну функцію — продукують секрет, який бере участь у регуляц водного обміну. Особливості у будові мають епендимоцити, що вкривають судинн сплетення шлуночків мозку та спинномозковий канал. Цитоплазма цих клітин на базальному полюсі має численні глибокі складки, значну кількість мітохондрій та включень. Ці епендимоцити беруть активну участь у синтезі цереброспінально рідини та регулюють її склад.
Астроцити утворюють опорний апарат центральної нервової системи. Це невеликі клітини зірчасто форми з численними відростками, які розходяться у різні боки. Розрізняють протоплазматичні та волокнисті (фібрилярні) астроцити, а також перехідні форми (волокнисто-протоплазматичні). Протоплазматичні астроцити локалізуються, переважно, у сірій речовині мозку, відростки їх короткі, товсті і дуже розгалужені. Волокнист астроцити, переважно, розмішуються у білій речовині мозку, відростки їх довгі, прямі, слабо розгалужені.
Відростки астроцитів закінчуються на судинах, нейронах, базальній мембрані, яка відокремлює мозкову тканину від м'язової оболонки. В усіх випадках відростки розширяються на кінц розплющуються на поверхні капіляра або нейрона, вкриваючи значну їх частину утворюючи так звану астроцитарну ніжку. Ніжки астроцитів контактують між собою формують майже повну обгортку навколо капіляра або нейрона.
У цитоплазмі астроцитів містяться фібрили, що складаються із філаментів. Кожний пучок зв'язу перинуклеарний простір з усіма відростками. Таким чином, цитоплазма астроцитів заповнена звивистими філаментами, які забезпечують міцність відростків, у ній мало елементів ендоплазматичного ретикулуму та рибосом. Популяція астроцитів може повільно відновлюватися.
Олігодендроцити — це найчисленніша група гліоцитів. Олігодендроцити оточують тіла нейронів та їхн відростки по всій довжині, локалізуються як у центральній, так і в периферійній нервовій системі. Вони характеризуються невеликими розмірами та дуже тонкими відростками. Перикаріони їх мають багатокутну або овальну форму. Цитоплазма олігодендроцитів не містить нейрофіламентів, судячи по швидкості асиміляц мічених атомів, у цих клітинах відбувається активний синтез білків і ліпідів. Олігодендроцити мають тісний контакт із структурами гемомікроциркулярного русла зв'язані із водним обміном мозку. При набряку мозку у їх відростках нагромаджується рідина. Функція олігодендроцитів дуже різноманітна: трофічна, золююча участь у водно-сольовому обміні, процесах дегенерації та регенерац нервових волокон. Олігодендроцити, які утворюють оболонки навколо відростків нервових волокон, називають або нейролемоцитами або швановськими клітинами.
Мікроглія — (гліальн макрофаги) — це спеціалізована система макрофагів, тканин внутрішнього середовища, які виконують захисну функцію. Це клітини з двома, трьома відростками, які мають вторинні розгалуження, ядра їх багаті на гетерохроматин. При подразненні нервової тканини (запалення, рана) клітини мікрогл збільшуються в об'ємі, стають рухомими, наповнюються фагоцитованим матеріалом. Мікрогліоцити здатні до синтезу білків-імуноглобулінів, вони характерні для білої і сірої речовини центральної нервової системи.
Таким чином, нервова тканина — складна система, яка складається з різноманітних нейронів та нейрогліальних елементів. Якщо не можна уявити собі нейрон поза рефлекторною дугою, ізольований від інших нейронів, оскільки ні рецепторний, н асоціативний, ні еферентний нейрони самі по собі не можуть виконати ніяко функції проведення нервового імпульсу, так і неможливо розглядати нервов клітини у відокремленні їх від нейроглії. Найбільш чітко проявляється взаємозв'язок нейронів і нейроглії при вивченні будови нервових волокон.
4. Нервові волокна
Нервові волокна — це провідники нервових імпульсів. Складаються вони із відростка нейрону — осьового циліндра і оболонки, що його вкриває (нейролемоцит, швановська клітина) структура олігодендроглії. Залежно від будови оболонки їх поділяють на дв основні групи — мієлінові та безмієлінові.
Безмієлінові нервов волокна є типовими для вегетативного відділу нервової системи, порівняно просто побудовані і спрямовуються, переважно, до внутрішніх органів. Нейролемоцити щільно прилягають один до одного, утворюючи безперервний тяж на поверхн нервового волокна. Складаються безмієлінові волокна з осьового циліндра, нейролеми і базальної мембрани. Прогинаючи оболонку нейролемоцитів, осьовий циліндр глибоко занурюється у цей тяж клітин, які, ніби муфта, огортають нервовий відросток. Входячи до лемоцита, осьові циліндри втягують за собою мембрану нейролемоцита і стають немов би підвішеними на подвійній складці мембрани, яку називають мезаксоном. В безмієліновому нервовому волокні міститься від 7 до 10 осьових циліндрів, які можуть переходити із одного безмієлінового волокна у сусіднє. У світловому мікроскопі ні мезаксона, ні межі між нейролемоцитами не видно, безмієлінове волокно здається суцільним пучком осьових циліндрів, обгороджених масою цитоплазми, яка містить ядра. Швидкість передачі нервового мпульсу у безмієлінових волокнах менша, ніж у мієлінових. Це пояснюється тим, що у безмієлінових волокнах хвиля деполяризації рухається по всій плазмолемі, не перериваючись. Відомо, що у внутрішніх органах рух, секреція та інші функц здійснюються повільніше, а тому осьові циліндри безмієлінових волокон не зольовані так чітко один від одного, як це має місце у мієлінових.
Мієлінові нервові волокна локалізуються як у центральній, так і у периферійній нервовій системі, тобто у складі головного та спинного мозку, а також периферійних нервах. Це товст волокна діаметром від 3 до 25 мкм, що складаються із осьового циліндра, мієлінової оболонки, нейролеми та базальної мембрани. Осьовий циліндр — це відросток нейрона, яким частіше буває аксон, але може бути і дендрит. Він складається з нейроплазми, яка містить поздовжньо орієнтовані нейрофіламенти нейротрубки, а також мітохондрії. Осьовий циліндр вкритий аксолемою (продовження плазмолеми нейрона), яка забезпечує проведення нервового імпульсу. В кожному мієліновому волокні лише один осьовий циліндр, навколо якого почергово, ланцюжком розміщуються нейролемоцити.
У процесі розвитку мієлінового волокна осьовий циліндр занурюється в нейролемоцит, втягуючи його плазмолему і утворюючи глибоку складку. Цю подвійну складку (дуплікатуру) нейролемоцита називають мезаксоном. У процесі подальшого ґенезу волокна, нейролемоцит (швановська клітина) повільно обертається навколо осьового циліндра, в результаті чого мезаксон багато разів огортає його.
Цитоплазма нейролемоцита його ядра залишаються на периферії, утворюючи нейролему волокна. Таким чином, мієлінова оболонка утворюється із щільно й концентрично нашарованих навколо осьового циліндра завитків мезаксона, які є пластинками мієлінового шару (рис. 1).
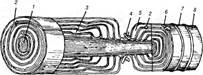
Рис.1. Схема будови мієлінового волокна:
1 — аксон; 2 — мезаксон; 3 — насічки; 4 — перехват; 5 — цитоплазма нейролемоцита; 6 — ядро нейролемоцита; 7 — невролема; 8 - ендоневрій
Кожний завиток мезаксона відповідає ліпідним шарам двох листків плазмолеми нейролемоцита. На його середині та поверхні на ультраструктурному рівні можна виявити темні лінії, утворені білковими молекулами. Насічки мієліну (Шмідта-Лантермана) відповідають тим місцям, де завитки мезаксону розсунуті цитоплазмою шванівської клітини. Насічки можуть мати напрямок як до центру, так і на периферію, у місцях насічок мієлін не переривається. Мієлінова оболонка волокна складається з багатьох нейролемоцитів, які, в місцях контакту між собою, утворюють перемички вузла (перехват Ранв'є). Переривність мієлінової оболонки має велике значення для транспортування до осьового циліндра необхідних речовин, сприяє обміну іонами та впливає на швидкість проведення нервового імпульсу.
Ділянка нервового волокна між двома вузловими перехватами називається міжвузловим сегментом, який відповідає одній гліальній клітині. На повздовжньому розрізі мієлінового волокна поблизу вузлового перехвату є ділянка, у якій завитки мезаксона послідовно контактують з осьовим циліндром. Місця прикріплення найглибших завитків найбільш віддалені від перетяжок, а всі наступні — поступово наближаються до них. Це пояснюється тим, що мезаксон нашаровується у процесі росту і осьового циліндра і нейролемоцитів. Таким чином, перші шари мезаксона коротші за останні. Краї двох сусідних лемоцитів, що контактують у ділянці перехвату Ранв'є, утворюють інтердигітації, які перекривають ділянку перемички і називаються «рихлим комірцем».
Мієлінові волокна центральної нервової системи у своїй будові мають ряд особливостей. їх оболонку утворюють типові олігодендроцити, у них відсутні інтердигітації, насічки Шмідта-Лантермана, відсутня сполучнотканинна базальна мембрана, роль яко виконує нейроглія.
Таким чином, у мієліновому волокні лише один осьовий циліндр, мезаксон, закручений щільними шарами, і утворює товстий шар мієліну. Порівняно великий діаметр осьового циліндра, наявність перехватів та добре розвинений мієліновий шар забезпечують швидке та точне проведення нервового імпульсу.
5. Синапси
Однобічна передача нервового імпульсу в межах рефлекторної дуги зумовлена спеціалізованим контактом двох нейронів, що називають міжнейронним синапсом. Морфологічно у складі синапсу розрізняють пресинаптичний і постсинаптичний полюси, між якими синаптична щілина. Зустрічаються синапси із хімічною та електричною передачами. Функціонально розрізняють два види синапсів — збудливі та гальмівні. Морфологічна класифікація їх залежить від того, які частини нейронів контактують між собою. Аксодендричні — аксон першого нейрона передає нервовий імпульс на дендрит другого. Аксосоматичні — аксон першого нейрона передає імпульс на перикаріон другого. Аксоаксонні — терміналі аксона першого нейрона закінчуються на аксоні другого. Очевидно, аксоаксонні синапси виконують гальмівну функцію. Крім останніх, найбільш поширених міжнейронних контактів, між деякими нейронами зустрічаються дендросоматичні та дендродендричні синапси. Таким чином, будь яка частина нейрона може утворювати міжнейронний синапс з будь-якою частиною іншого нейрона.
Морфологічно пресинаптичний полюс синапса утворений термінальною гілочкою аксона т нервової клітини, яка передає імпульс. Цей полюс, здебільшого, розширений у вигляді ґудзика, вкритий пресинаптичною мембраною. У ньому містяться мітохондрії та синаптичні пухирці, вкриті мембраною і з вмістом певних хімічних речовин, так звані медіатори. Останні сприяють передачі нервового імпульсу на постсинаптичну частину міжнеиронного контакту. Синаптичні пухирці бувають різних розмірів (маленькі прозорі, великі електронно-щільні, прозорі, що містять щільну гранулу). Вони містять хімічні речовини, що називаються медіаторами. Участь в передачі нервового імпульсу іншому нейрону відбувається за рахунок транспортування шляхом екзоцитозу в синаптичну щілину медіатора. Медіаторами можуть бути ацетілхолін (холінергічні синапси), норадреналін та адреналін (адренергічні синапси), а також інші речовини — серотонін, глутамінова кислота, нейротензин, ангіотензин, речовина Р, енкефалін (у збудливих синапсах) дофамін, гліцин, гамма-аміномасляна кислота (є медіаторами гальмівних синапсів).
Пресинаптична мембрана містить електронно-щільні частинки, які разом із нейрофіламентами утворюють пресинаптичну решітку для пухирців. У холінергічних синапсах ці пухирці дрібн прозорі, а також зустрічаються великі та електронно-щільні, синаптичн пухирці адренергічних синапсів великі, прозорі і морфологічно характеризуються наявністю електронно-щільної гранули.
Постсинаптична мембрана холінергічних синапсів містить «холінрецептор-ний» білок — рецептор медіатора, чим зумовлена дія останнього на постсинап-тичну мембрану. При взаємод рецептора медіатора з ацетилхоліном утворюють конформаційні зміни його молекул, які призводять до зміни проникності мембрани та генерації нервового імпульсу в нейроні. Медіатори гальмівних синапсів (дофамін, гліцин) не збільшують проникність мембрани для іонів, а зменшують її, стабілізуючи мембранний потенціал, а, отже, гальмують генерацію нервового імпульсу.
Синаптична щілина заповнена тканинною рідиною, в якій містяться електронно-щільні частинки та ниткоподібні структури, що розміщуються на поверхнях обох синаптичних мембран. Очевидно, ці структури містять одночасно пре- і постсинаптичні мембрани. При проходженні нервового імпульсу до закінчення пресинаптичного нейрона синаптичн пухирці зливаються із пресинаптичною мембраною, їхній вміст виливається в щілину, і медіатор діє на постсинаптичний нейрон. Мембрана пухирців використовується повторно.
Електротонічні синапси утворюються при щільному приляганні нейролем двох нейронів, переважно, їх дендритів і перикаріона. Це, так звані, закриті, електричні синапси (безпухирцеві). Останні не мають синаптичної щілини.
6. Нервові закінчення
Термінальні апарати нервових волокон — нервові закінчення за їх функціональним значенням поділяють на три види: рецептори, ефектори та міжнейронні синапси.
Рецептори (чутлив нервові закінчення) — спеціалізовані термінальні структури дендритів нейронів, пристосовані до сприйняття подразнень, що надходять до організму. Залежно від локалізації і специфічності нервової регуляції життєдіяльності організму рецептори поділяють на екстерорецептори (сприйняття подразнень із зовнішнього середовища) та інтерорецептори (подразнення власних тканин організму). Різновидом інтерорецепторів пропріорецептори — нервові закінчення у м'язах та сухожиллях, які беруть участь у регуляції рухів і положенні тіла у просторі. Залежно від природи подразнень, останні поділяють на термо-, механо-, баро-, хемо-, ноціорецептори та інш.
Чутливі нервов закінчення надзвичайно різноманітні за структурною організацією. Вони поділяються на вільні нервові закінчення, які складаються лише із термінальних розгалужень осьового циліндра дендрита чутливого нейрона та невільні — крім осьового циліндра, містять також клітини нейроглії. Якщо невільні рецептори оточені сполучнотканинною капсулою, їх називають капсульованими; ті невільн рецептори, що не мають сполучнотканинної капсули, мають назву некапсульовані. Рецепторні закінчення або рецепторно-нейтральні синапси складаються із спеціалізованих клітин, здатних реагувати на подразник і викликати деполяризацію плазмолеми зв'язаного з ним дендрита аферентного нейрона.
Прикладом вільного рецептора можуть бути деревоподібні та кущеподібні нервові закінчення епітеліальної тканини. При формуванні таких закінчень, мієлінові нервові волокна, підходячи до епітеліального пласта, втрачають мієлінову оболонку, а їхні осьов циліндри утворюють кінцеві розгалуження, що розміщуються між окремими епітеліоцитами. За функцією це, як правило, термо- та ноцірецептори. Вільні нервові закінчення можуть обплітати у вигляді кошика волосяні фолікули, реєструючи зміщення у просторі окремих волосин, відіграючи роль механорецепторів.
До складу рецепторного апарату багатошарових епітелієв належать поодинокі чутливі епітеліальн клітини, так звані дотикові епітеліоцити Меркеля. Ці електронно-прозорі клітини мають сплющені ядра та осьміофільні гранули. До їх базальної частини прилягають чутливі нервові закінчення у вигляді дисків (дотикові меніски Меркеля). Нейроепітеліальн клітини Меркеля мають постійне місце в епітеліальному шарі, незважаючи на постійний рух епітеліоцитів від базального до дистального полюса, утворюючи шари епітелію. Клітини Меркеля — типовий приклад нейроепітелію, епітеліоцити якого пристосувалися до рецепції, вони виконують функцію механорецепції.
Надзвичайно різноманітн рецепторні апарати сполучної тканини представлені двома групами невільних нервових закінчень. У капсульованих рецепторах нервові терміналі аферентних волокон, як правило, оточені нейролемоцитами і допоміжними елементами сполучнотканинного походження. Серед капсульованих чутливих закінчень, залежно від будови, розрізняють тільця Фатер-Пачіні, Гольджі-Маццоні, Мейснера, кінцеві колби Краузе (генітальні тільця). Найбільш поширені рецептори сполучної тканини — це пластинчасті тільця Фатер-Пачіні. У їх складі розрізняють внутрішню колбу та зовнішню капсулу. Навколо розгалужень осьового циліндра (термінальний відділ чутливого нервового волокна), яке втратило мієлінову оболонку, скупчуються видозмінені нейролемоцити, що і утворюють внутршіню колбу. Капсула тілець Фатер-Пачіні складається із великої кількості сполучнотканинних пластинок, утворених фібробластами та спірально орієнтованими пучками колагенових волокон. На межі внутрішньої колби та зовнішньої капсули розмішуються клітини, які, очевидно, визначаються як гліальні. Ці гліоцити утворюють синапси із розгалуженням осьового циліндра. Є припущення, що нервовий імпульс генерується в умовах зміщення зовнішньої капсули щодо внутрішньої колби.
Тільця Гольджі-Маццон менші від тілець Фатер-Пачіні, мають тонку капсулу і, порівняно, велику внутрішню колбу. Ці структури виконують функцію барорецепції у шкірі, серозних та слизових оболонках.
Тільця Мейснера (дотикові) — це рецептори тактильної чутливості, які локалізуються у сосочковому шарі дерми. Ці структури орієнтовані перпендикулярно до поверхн шкіри і мієлінові волокна, втрачаючи мієлінову оболонку, контактують із поверхнею гліальних клітин колби. Колагенові волокна сполучнотканинної капсули можуть заповнювати простори між нейролемоцитами і терміналями чутливого нервового волокна.
Кінцеві колби Краузе механо- і терморецепторами кон'юнктиви, язика, а видозмінені структури (генітальні тільця) — зовнішніх статевих органів. Характерною особливістю будови колб Краузе є дуже тонка сполучнотканинна капсула. Мієлінове волокно, входячи в капсулу, втрачає мієлінову оболонку і закінчується колбоподібним розширенням. Генітальні тільця мають також таку будову, за винятком того, що від капсули проникає не один осьовий циліндр, а декілька, які, розгалужуючись, утворюють сіточку безмієлінових волокон.
Нервово-сухожильн веретена (сухожильні органи Гольджі) утворені товстими мієліновими волокнами, які, підходячи до колагенових волокон, втрачають мієлінову оболонку і утворюють численні розгалуження, обплітають сухожильні пучки. Подібну будову мають тільця Руффіні, що локалізуються у глибоких шарах дерми та підшкірній жировій тканині. Нервово-сухожильні веретена, як і тільця Руффіні вважають механорецепторами, які сприймають взаємне зміщення колагенових волокон.
Нервово-м'язові веретена це чутливі нервові закінчення поперечно-смугастої м'язової тканини. Вони здатні сприймати зміну довжини м'язового волокна і швидкість цієї зміни. Нервово-м язові веретена містять кілька інтрафузальних м'язових волокон, вкритих загальною сполучнотканинною капсулою. До складу веретена входять два товстих центральних м'язових волокна та чотири тонких. Екваторіальна частина товстих волокон заповнена скупченням ядер — «ядерна сумка». Нервово-м'язове веретено має два типи нервових волокон. Одні утворюють спіральні закрутки, що оточують екваторіальну частину товстих інтра-фузальних (внутрішньо-веретенних) м'язових волокон — це кільцево-спіральні «анулярні» закінчення. Друга група — закінчення чутливих волокон, в своєму складі містить як кільцево-спіральні, так гроноподібні терміналі, що розміщуються по одному з кожного боку кільцеподібних. Останні реагують на зміну довжини м'язового волокна та швидкість зміни, гроноподібні — лише на зміну довжини м'язового волокна. На обох полюсах м'язових волокон локалізуються терміналі рухових нервових волокон.
Ефектори — структури, утворені закінченням аксонів нейроцитів, які бувають рухові та секреторні. Нервово-м'язові закінчення у скелетних м'язах утворені терміналями аксонів нейроцитів рухових ядер спинного та моторних — головного мозку. При наближенн до м'язового волокна мієлінове волокно втрачає мієлінову оболонку, його осьовий циліндр розгалужується і разом із саркоплазмою міосимпласта занурюється у м'язове волокно. У цій ділянці утворюється аксом'язовий синапс, де аксолема відіграє роль пресинаптичної мембрани, а сарколема м'язового волокна постсинаптичною мембраною. Медіатор цих синапсів — ацетилхолін, який при збудженні вивільняється і через пресинаптичну мембрану проникає у синаптичну щілину. На поверхні постсинаптичної мембрани ацетилхолін зв'язується із холінорецепторами і стає хімічним сигналом для збудження м'язового волокна.
Еферентні закінчення у гладенькій м'язовій тканині мають простішу будову. Окремі нервові закінчення на поверхні гладеньких міоцитів утворюють варикозні терміналі. Медіатор цих закінчень — ацетилхолін, або адреналін.
Вивчаючи клітини тканин, ми постійно користувалися методом аналізу, виділення із організму його окремих частин. В подальшому нам необхідно, використовуючи ці аналітичні дані для розуміння того, як із вивчених нами тканин будуються органи, системи органів увесь організм в цілому, що розглядається у розділі спеціальної гістології.
Література
1. Быков В.Я. Цитология. Общая гистология. - СПб, 1999.
2. Гистология, цитология и эмбриология / Под ред. О.В. Волкова, Ю.К. Елецкого. - М: Медицина, 1996.
3. Новак В.П., Пилипенко М.Ю., Бичков Ю.П. Цитологія, гістологія, ембріологія.: Підручник. – К.: ВІРА-Р, 2001. – 288 с.
© 2009 База Рефератов